Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.___________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ___________________ 205
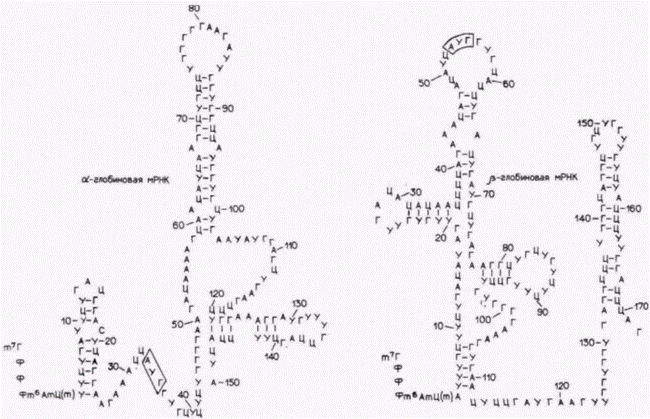
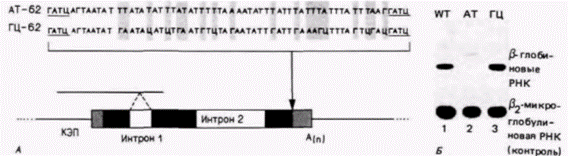
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.206______________ ГЛАВА 14_____________________________________________________________________________ водится с одной молекулы мРНК фактора роста. Основной элемент, контролирующий время жизни мРНК, находится, по-видимому, в 3'-нетранслируемой области мРНК. При этом молекулы эпидермальной РНК содержат одну или несколько АУ-богатых последовательностей в этой области. В одном из опытов АТ-богатый участок длиной в 51 пару оснований из 3'-нетранслируемой области гена для фактора роста КСФ-г,м был встроен в 3'-нетранслируемую область ß-глобинового гена кролика (рис. 14.10; Shaw, Kamen, 1986). Время полужизни полученной глобиновой мРНК составляло менее 30 мин. Аналогичная последовательность, но содержащая 14 остатков Г и Ц, была встроена в качестве контроля в другой ß-глобиновый ген. Соответствующая глобиновая мРНК имела обычное время полужизни. Создастся впечатление, что продолжительность существования конкретной мРНК закодирована в 3'-нетранслируемой области этой мРНК. Способность к дифференциальной деградации различных мРНК имеет критическое значение для клеточных функций. Например, ген c-fos кодирует ядерный белок, необходимый для деления нормальных фибробластов (Holt et al., 1986). Подобно мРНК фактора роста КСФ-г,м, мРНК для c-fos имеет протяженную 3'-нетранслируемую область, богатую АУ-последовательностями. Если эта область делетирована (экспериментально или при спонтанных мутациях), то время полужизни соответствующей мРНК увеличивается. Следовательно, производится больше белка c-fos и клетка непрерывно стимулируется к делению. В результате возникает опухоль из клеток, в которых мРНК гена c-foc лишена АУ-богатой 3'-области (Meijlink et al., 1985). Трансляционный контроль ооцитных мРНКДанные, свидетельствующие о материнской регуляции раннего развитияУ большинства видов животных экспрессия генов диплоидного ядра происходит не сразу. Данные о том, что раннее развитие контролируется факторами, запасенными или синтезированными в ооците, получены в ряде экспериментов на рубеже нашего века (см. обзор Davidson, 1976). Результаты этих экспериментов отчетливо показали доминантность материнских признаков на начальных стадиях эмбриогенеза и переключение на отцовские или гибридные характеристики только в более позднем развитии. О таких долговременных материнских влияниях мы упоминали при обсуждении ориентации дроблений у зародышей улитки, у которых Таблица 14.2. Данные Дриша по материнскому контролю числа клеток первичной мезенхимы в гибридных зародышах морского ежа (Источник: Davidson, 1976.)
цитоплазма ооцита содержит фактор, управляющий поворотами плоскостей дробления в правом или левом направлении. В 1898 г. Ханс Дриш скрестил два вида морских ежей и обнаружил, что среднее число первичных клеток мезенхимы в гибриде зависит исключительно от вида, давшего яйцо (табл. 14.2). В сходном эксперименте яйцеклетки морского ежа Cidaris оплодотворяли спермой морского ежа Lytechinus (Tennant, 1914). При этом было обнаружено, что в образующихся бластомерах сохраняются все хромосомы, а формирование архентерона и время образования мезенхимы соответствуют материнскому типу развития (табл. 14.3). Отцовские гены проявлялись впервые при закладке скелетных клеток. Таблица 14.3 Данные Теннанта по контролю этапов ранней гаструляции в гибридных зародышах морского ежа
| ||||||||||||||||||||||