Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

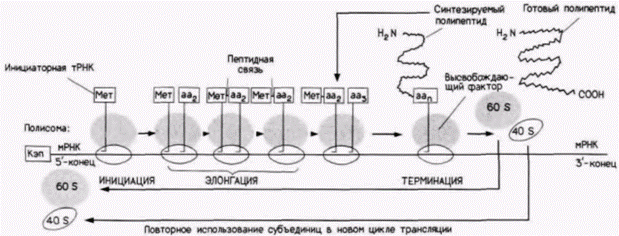
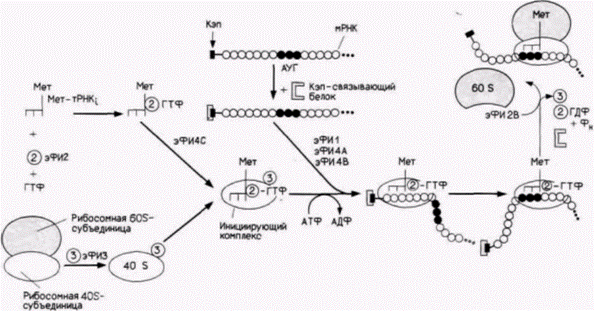
Глава 14. Трансляционная и посттрансляционная регуляция процессов развитияМы не должны игнорировать тот факт, что каузальное исследование организмов является одной из наиболее сложных, если не самой сложной проблемой, которую пытался решить человеческий интеллект, и что это исследование, подобно любой другой каузальной науке, никогда не может быть завершено, так как каждая вновь установленная причина только поднимает новые вопросы о причине данной причины, ВИЛЬГЕЛЬМ РУ (1894) У посыльного нет отдыха до тех пор. пока послание не вручено. ДЖОЗЕФ КОНРАД (1920) ТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯПосле транскрипции, процессинга и выхода мРНК из ядра необходима ее трансляция, чтобы получить белок, закодированный в геноме. Из этой главы мы узнаем, что регуляция на уровне трансляции представляет собой исключительно важный механизм контроля экспрессии генов. В данном случае мРНК уже образована, но она может транслироваться или не транслироваться в зависимости от определенных условий в клетке. Таким образом, контроль экспрессии генов на уровне трансляции может использоваться, когда нужен немедленный всплеск синтеза белка (как мы увидим, это происходит сразу после оплодотворения яйца) или в качестве механизма тонкой регуляции, обеспечивающей синтез строго определенного количества белка на имеющемся запасе мРНК (например, при синтезе гемоглобина). Мы узнаем также, что контроль на уровне трансляции осуществляется несколькими путями и в различных клетках используются разные пути. Механизм трансляции у эукариотТрансляция – это процесс, с помощью которого информация, содержащаяся в нуклеотидной последовательности мРНК. управляет синтезом конкретного полипептида. Этот процесс можно разделить на три этапа: инициацию, элонгацию и терминацию (рис. 14.1). На первом этапе инициации – происходит присоединение первой аминоацил-транспортной РНК и мРНК к рибосоме. Единственной транспортной РНК (тРНК), способной к инициации трансляции, является особая тРНК (тРНК,), которая несет аминокислоту метионин. Как показано на рис. 14.2, первые реакции вызывают образование инициирующего комплекса, состоящего из метиониновой инициаторной тРНК, связанной с 40S(«малой»)-субъединицей рибосомы. Эта Мет-тРНК, узнается эукариотическим фактором инициации 2 (эФИ2), который присоединяет ее к 40S-субъединице рибосомы. Отметим, что присоединение происходит в отсутствие мРНК. Следующей добавляется мРНК. Кэп-связывающий белок присоединяется к 7-метилгуанозину кэп-группы на 5'-конце мРНК, а эукариотические факторы инициации 4А и 4В прикрепляются к кэп-связывающему белку или рядом с ним. В отсутствие кэп-группы связывание мРНК с рибосомной субъединицей неполноценное (Shatkin, 1976, 1985). Рибосомная 40S-субъединица перемещается вдоль мРНК, пока не достигает кодона АУГ в правильном окружении. Было показано, что не каждый АУГ-кодон подходит для этого (Kozak, 1986). Для того чтобы остановить рибосомную 40S-субъединицу и инициировать трансляцию, важны также нуклеотиды вокруг АУГ. С помощью мутирования клонированных генов и анализа трансляции их РНК было обнаружено, что «оптимальной» является последо· Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.200_______________ ГЛАВА 14_____________________________________________________________________________
вательность АЦЦАУГТ. Мутации среди фланкирующих нуклеотидов могут снижать трансляцию в 20 раз. Важность фланкирующей последовательности наблюдалась также in vivo. Имеется сообщение (Morle et al., 1985) о больном α-талассемией (дефицит α-глобиновых субъединиц), вызванной изменением последовательности АЦЦАУГГ на ЦЦЦАУГГ. При связывании 40S-субъединицы с АУГ в мРНК инициаторная тРНК располагается над АУГ-кодоном. Только после правильного размещения мРНК на малой рибосомной субъединице может присоединиться рибосомная («большая») 60S-субъединица.
|