Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ___________________ 201
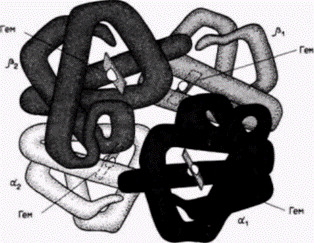
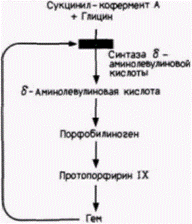
Элонгация заключается в последовательном присоединении молекул аминоацил-тРНК к рибосоме и образовании пептидных связей между аминокислотами по мере того, как они последовательно отдают свои тРНК-переносчики (рис. 14.1). После соединения аминокислот друг с другом рибосома перемещается по мРНК, экспонируя новые кодоны для связывания тРНК. Это позволяет другой рибосоме инициировать трансляцию на 5'-конце мРНК и начать свое перемещение. Таким образом, с любой мРНК связано обычно несколько рибосом. Эту структуру называют поэтому полирибосомой или, что более принято, полисомой (рис. 14.3). Терминация синтеза белка происходит, когда на рибосоме экспонируется один из кодонов мРНК УАГ, УАА или УГА. Эти триплеты нуклеотидов (называемые кодонами терминации) не узнаются молекулами тРНК и, следовательно, не кодируют ни одной аминокислоты. Однако их узнают сбрасывающие факторы, которые гидролизуют связь пептида с последней тРНК, освобождая его от рибосомы. Рибосома разделяется на две субъединицы, и цикл трансляции начинается снова. Хотя 3'-поли(А)-хвосты молекул мРНК не транс- Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.202_______________ ГЛАВА 14_____________________________________________________________________ лируются, они, по-видимому, увеличивают эффективность трансляции тех мРНК, у которых они имеются. Как правило, пока мРНК находится в цитоплазме, ее поли(А)-хвост постепенно укорачивается, и если размеры его станут ниже критического, то способность мРНК к трансляции существенно снижается (Littauer, Soreq, 1982). В некоторых случаях экспрессия различных мРНК в цитоплазме может регулироваться с помощью дифференциального укорачивания поли(А)-хвостов. Этот механизм наблюдали в слюнных железах личинки Drosophila (Restifo, Guild, 1986) и в ооцитах Xenopus(Dworkin, Dworkin-Rastl, 1985). У миксомицета Dictyostelium(развитие которого рассмотрено в гл. 1) и двустворчатого моллюска Spisula дифференциальное укорачивание поли(А)-хвостов играет решающую роль в их жизненном цикле. При переходе миксомицета от вегетативного роста (амеба) к развитию (плодовое тело) транскрибируется новый набор мРНК. Одновременно резко укорачиваются поли(А)-хвосты запасенных мРНК, специфичных для вегетативной стадии. В результате новосинтезированные мРНК транслируются, а запасенные мРНК нет. Высказано предположение, что поли(А)-хвост каким-то образом поддерживает трансляцию (Palatnik et al., 1984). Эта идея согласуется с наблюдениями, что добавленная экзогенная поли(А) ингибирует трансляцию мРНК с такими хвостами (Jacobson, Favreau, 1983). Сходным образом после оплодотворения ооцита Spisula поли(А)-хвосты в молекулах мРНК, транслируемых в ооците, сильно укорачиваются, тогда как в тех мРНК, которые будут транслироваться у ранних зародышей (и которые не транслировались в ооците), удлиняются (Rosenthal, Ruderman, 1987). Контроль на уровне трансляции при координированном синтезе белка: продукция гемоглобинаОдной из основных проблем генетической регуляции является координированный синтез нескольких продуктов с разных участков генома. Когда развивающийся эритроцит синтезирует гемоглобин, необходимо, чтобы α-глобиновые цепи, ß-глобиновые цепи и молекулы гема производились соответственно в соотношении 2:2:4 (рис. 14.4). Любое существенное отклонение от этого соотношения приводит к тяжелым заболеваниям. Результаты недавних исследований показали, что пропорциональный синтез компонентов гемоглобина регулирует молекула гема. Достигается это двояким образом. Во-первых, избыток гема (т.е. гема, который не связан с белком, подобным глобину) будет выключать свой собственный синтез (Karibian, London, 1965). Осуществляется выключение
синтеза посредством инактивации 6-аминолевулинатсинтазы (DALA-синтазы), первого фермента на пути продукции гема (рис. 14.5). Таким образом, когда количество гема превосходит количество молекул, способных присоединить его, дальнейшая продукция гема прекращается. Во-вторых, избыток гема стимулирует синтез глобинов (Gribble, Schwartz, 1965; Zucker, Schulman, 1968). Если гем (в виде своей окисленной формы – гемина) добавляют к бесклеточной системе трансляции, в состав которой входят факторы, необходимые для трансляции мРНК (табл. 14.1), то синтез глобина значительно увеличивается (рис. 14.6). Следовательно, если для связывания гема не хватает глобинов, то избыток гема выключает свой собственный синтез и стимулирует усиленную продукцию глобинов. |