|
|
Ответы на билеты по биологии. Билет1 Закономерности существования клетки во времени. Жизненный цикл клетки, его варианты. Основное содержание и значение периодов жизненного цикла клетки
Диагноз: тениоз.
Возбудитель тениоза — Taenia solium (Linnaeus, 1758) рода Taenia, свиной, или вооруженный, цепень — ленточный гельминт, длиной 1,5—2 м (иногда до 3—4 м). Его стробила содержит до 1000 члеников. Сколекс 1—2 мм в диаметре вооружен 4 присосками и венчиком хитиновых крючьев.
Молодые членики — бесполые, средней трети — гермафродитные, зрелые (размер—12— 15X6—7 мм) содержат матку с боковыми ответвлениями от 6 до 12 пар Тонкие членики неподвижны и, отторгаясь от стробилы, выделяются во внешнюю среду при дефекации. В каждом членике содержится от 30 до 50 000 яиц. Яйца и онкосферы свиного и бычьего цепня морфологически одинаковы.
Окончательный хозяин возбудителя тениоза — человек, промежуточный облигатный — свинья. Факультативными промежуточными хозяевами являются человек, собаки, кошки, обезьяны. У окончательного хозяина гельминты паразитируют в тонкой кишке. Членики выделяются с испражнениями, при их разрушении освобождаются яйца, онкосферы, которыми при поедании нечистот заражаются животные. Освободившиеся в кишках животного зародыши проникают в ток крови и заносятся в межмышечную соединительную ткань, где спустя 2—3 мес образуются финны. Человек заражается при употреблении свиного финнозного мяса. Полное развитие Т. solium завершается через 2,5 — 3 мес.
Патогенез, клиника тениоза. В патогенезе тениоза выражено механическое, нервно-рефлекторное и аллерго-токсическое действие гельминтов.
У больных наблюдаются расстройства функции кишечника: тошнота, рвота, иногда понос, схваткообразные боли в животе, потеря веса; Возможно развитие холецистита, панкреатита. Больных беспокоит слабость, головокружение, периодические головные боли, нарушения сна. Иногда тениоз сопровождается анемией. Нередко клинические проявления отсутствуют.
Профилактика. Мясо и мясные продукты следует приобретать только в магазинах, а не на стихийных рынках:
- при покупке мяса на рынке следует требовать наличие документа о проведенной ветеринарно-санитарной экспертизе и проверять, имеется ли на мясе клеймо;
- употреблять в пищу следует только хорошо проваренное и прожаренное мясо;
- в процессе приготовления мясных блюд не пробовать сырой фарш.
Билет №18.
1).Ген как единица изменчивости. Генные мутации и их классификация. Причины и механизмы возникновения генных мутаций. Мутон. Последствия генных мутаций для человека.
Ген-это единица наследственности и изменчивости. Каждый ген распологается в определенном участке хромосомы-локусе. Два гена находящиеся в одном локусе называются алельными. Изменчивость-свойство живых организмов приобретать в процессе онтогенеза отличия от родительских форм. Различают фенотипическую (не наследственную) и генотипическую(наследственную) изменчивость. Фенотипическая изменчивость не связанна с изменением генотипа и не сохраняется в следующем поколении, если нет условий, в которых она возникла(изменяется лишь функциональная активность генов).
Фенотипическая изменчивость бывает возрастная и модификационная.
Возрастная, или онтогенетическая выражается в изменении всего комплекса морфофизиологических и биохимических признаков организма на протяжении его индивидуальной жизни.
Модификационная - возникает у генетически однородных особей одинакового возраста, которые находятся в разных условиях внешней среды, и проявляется в организме в качественных и количественных отклонениях. Диапазон модификационной изменчивости, в рамках которой зависимо от условий внеш.среды один и тот же генотип способен давать разные фенотипы, называют нормой реакции.
Комбинативная изменчивость-связанна с новыми комбинациями генов в генотипе. Возникает такая изменчивость в результате процессов:
1.Независимого расхождения хромосом в мейозе( особый вид деления клеток, в результате которого образуются гаметы - половые клетки)
2.Случайного сочетания генов при оплодотворении
3.Рекомбинации генов благодаря перекрещиванию хромосом.
Мутационная изменчивость- скачкообразное и устойчивое изменение генетического материала,передающегося по наследству. Возникает в следствии резких изменений признаков-мутаций и обусловливает реорганизацию структур воспроизведения, перестройку генетического аппарата. Этим мутации резко отличаются от модификации,которые не зацапают генотип особей. Мутации возникают внезапно, скачкообразно.образуя иногда формы организма , резко отменные от исходной формы.
1. Изменчивость, ее причины и методы изучения. Классификация форм изменчивости. Фенотипическая изменчивость и ее компоненты. Наследуемость признаков
Самовоспроизведение с изменением – это одно из основных свойств жизни. Термин «изменчивость» служит для обозначения различных понятий; как и большинство других терминов, он полисемантичен (многозначен). Юрий Александрович Филипченко различал два основных подхода к определению изменчивости.
1. Изменчивость как состояние. В этом значении термин «изменчивость» служит для обозначения отличий биологических объектов друг от друга в данный момент времени. Всегда существуют различия между частями одного организма, между разными организмами в популяции, между разными внутрипопуляционными группировками, между популяциями.
2. Изменчивость как процесс. В этом значении термин «изменчивость» служит для обозначения изменения биологического объекта во времени. В этом случае изменчивость отражает развитие особи, отличие потомков от родителей.
Любая наблюдаемая изменчивость является фенотипической. В свою очередь, фенотипическая, или общая изменчивость включает три компонента:
– Наследственная (генетическая, или генотипическая изменчивость) – в значительной мере обусловлена влиянием генетических факторов. Например, в сходных условиях выращивается несколько сортов одного вида растений. Тогда различия между результатами эксперимента (например, урожайность) обусловлены генетическими особенностями каждого сорта. В основе генетической изменчивости лежит мутационная и комбинативная изменчивость.
– Ненаследственная (модификационная) изменчивость – в значительной мере обусловлена действием негенетических (экзогенных) факторов. Например, один сорт растений выращивается в разных условиях. Тогда различия между результатами эксперимента (например, урожайность) обусловлены влиянием условий выращивания растений.
– Неконтролируемая (остаточная изменчивость) – обусловлена неконтролируемыми (по крайней мере, в данном эксперименте) факторами.
Для разных признаков влияние генотипа и условий среды на общую фенотипическую изменчивость неодинаково. Например, окраска шерсти, жирномолочность у крупного рогатого скота, масса яиц у кур зависят, в основном, от особенностей породы (т.е. от генотипа) – эти признаки обладают высокой наследуемостью. Другие признаки: качество шерсти, общая удойность у КРС, яйценоскость у кур – зависят, в основном, от условий выращивания и содержания – эти признаки обладают низкой наследуемостью.
Генные мутации. Последствия мутаций.
Генные мутации выражаются в изменении структуры отдельных участков ДНК. По своим последствиям генные мутации делятся на две группы: мутации без сдвига рамки считывания и мутации со сдвигом рамки считывания.
Мутации без сдвига рамки считывания происходят в результате замены нуклеотидных пар, при этом общая длина ДНК не изменяется. В результате возможна замена аминокислот, однако из-за вырожденности генетического кода возможно и сохранение структуры белка.
Пример 1. Замена аминокислотного остатка в составе полипептида (миссенс–мутации). В состав молекулы гемоглобина человека входят две a-цепи (a-цепь закодирована в 16-ой хромосоме) и две b-цепи (b-цепь закодирована в 11-ой хромосоме). В состав b-цепи входит 146 аминокислотных остатков, при этом в нормальной b–цепи шестым аминокислотным остатком является глутаминовая кислота. С участием нормальной b-цепи образуется нормальный гемоглобин – HbA. В нетранскрибируемой нити участка ДНК, кодирующего b-цепь, глутаминоваякислота закодирована триплетом ГАА. Если же в результате мутации в ДНК произойдет замена триплета ГАА на триплет ГТА, то на местеглутаминовой кислоты в молекуле гемоглобина в соответствии с генетическим кодом появится валин. В итоге вместо гемоглобина HbAпоявится новый гемоглобин – HbS. Такая замена всего лишь одного нуклеотида и одной аминокислоты приводит к развитию тяжелого заболевания – серповидноклеточной анемии.
На клеточном уровне серповидноклеточная анемия проявляется в том, что при гипоксии (недостатке кислорода) эритроциты приобретают форму серпа и теряют способность к нормальному транспорту кислорода. Гомозиготы HbS/HbS умирают в раннем детстве. Зато гетерозиготы HbA/HbS характеризуются слабо измененными эритроцитами. При этом изменение формы эритроцитов значительно повышает устойчивость гетерозигот к малярии. Поэтому в тех регионах Земли, где свирепствует малярия (например, в Африке), отбор действовал в пользу гетерозигот. Таким образом, серповидноклеточная анемия – это пример относительности «полезности» и «вредности» мутаций.
Пример 2. Мутация без замены аминокислотного остатка в составе полипептида (сеймсенс-мутации). Если в нетранскрибируемой нити участка ДНК кодирующего b–цепь гемоглобина, произойдет замена триплета ГАА на триплет ГАГ, то из-за избыточности генетического кода замены глутаминовой кислоты не произойдет. В итоге структура b–цепи гемоглобина не изменится, и в эритроцитах будет обнаруживаться только нормальный гемоглобин HbA. Таким образом, вовсе не любая генная мутация проявляется в фенотипе.
Особую группу образуют ликовые мутации, в результате которых происходит незначительное изменение характеристик конечного продукта. Это связано с заменой аминокислотных остатков в пассивной части белка: такие замены не оказывают существенного влияния на структуру и функции белка.
Мутации со сдвигом рамки считывания (фреймшифты) происходят в результате вставки или потери нуклеотидных пар, при этом общая длина ДНК изменяется. В результате происходит полное изменение структуры белка.
Однако если после вставки пары нуклеотидов происходит потеря пары нуклеотидов (или наоборот), то аминокислотный состав белков может восстановиться. Тогда две мутации хотя бы частично компенсируют друг друга. Это явление называется внутригенной супрессией.
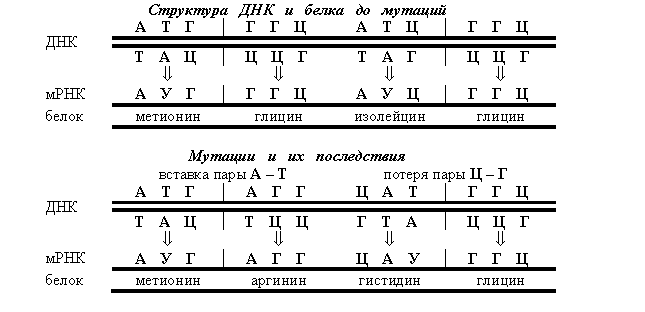
Мутации со сдвигом рамки считывания составляют
80% от всех генных мутаций. Вставки иначе называются инсерциями, а потери –эксцизиями. Процесс образования вставок называется инсерционным мутагенезом. Инсерционный мутагенез необходимо учитывать в генной инженерии.
Нонсенс–мутации. Особую группу генных мутаций составляют нонсенс-мутации с появлением стоп-кодонов (замена смыслового кодона настоп-кодон). Нонсенс-мутации могут возникать как вследствие замен нуклеотидных пар, так и с потерями или вставками. С появление стоп-кодонов синтез полипептида вообще обрывается. В результате могут возникать нуль-аллели, которым не соответствует ни один белок. Соответственно, возможно и обратное явление: замена нонсенс-кодона на смысловой кодон. Тогда длина полипептида может увеличиваться.
Дополнение 1. Существуют особые мутации, влияющие на экспрессию генов у эукариот
1. Мутации, изменяющие степень компактизации ДНК. В гигантских политенных хромосомах и в хромосомах типа ламповых щеток описаны мутации, инактивирующие ген, расположенный в каком-либо одном участке ДНК, т.е. блокирующие декомпактизацию хроматина. Скрещивание гетерозигот по таким регуляторным мутациям в F2дает расщепление 3:1, указывая на то, что они затрагивают единичные менделирующие факторы.
2. Гомеозисные мутации. Изменяют порядок экспрессии генов. Фенотипический эффект гомеозисных мутаций заключается в превращении одних органов в другие.Например, у мушки дрозофилы мутация группы bithorax, контролирующих развитие грудных и брюшных сегментов у дрозофилы, может приводить к появлениюкрылоподобных образований вместо галтеров; мутации группы antennapedia выражаются в том, что у насекомых на месте антенн вырастают ножки; мутацииophthalmoptera – развитие крыла из имагинального диска глаза; мутации proboscipedia – развитие ноги или части антенны (в зависимости от температуры) вместо хоботка;у мутантов tumorous head ткани головы замещаются другими типами тканей, включая структуры, характерные для гениталий.
Дополнение 2. Некоторые мутации обладают плейотропным действием, т.е. приводят к изменению сразу нескольких признаков.
Пример 1. Ароматические аминокислоты – триптофан, фенилаланин, тирозин – образуются из хоризмовой кислоты. Если некоторая мутация заблокирует хотя бы один этап синтеза хоризмовой кислоты, то клетка (организм) утрачивает способность к синтезу сразу трех аминокислот.
Пример 2. Один и тот же фермент (трансаминаза) контролирует синтез валина (из α–кетоизовалериановой кислоты) и изолейцина (из α–кето–β–метилвалериановой кислоты). Если некоторая мутация нарушит функции этого фермента, то клетка (организм) утрачивает способность к синтезу сразу двух аминокислот.
Пример 3. Один и тот же полипептид (продукт экспрессии одного гена) может входить в состав разных ферментов. Например, белок-апофермент липоатдегидрогеназыкишечной палочки в качестве субъединицы входит в состав других ферментов: пируватдегидрогеназы, 2-оксоглутаратдегидрогеназы, глицинового расщепляющего комплекса. Тогда мутация в гене LDH скажется на активности всех перечисленных ферментов.
Дополнение 3. Мутация в одном гене может подавлять мутации, происходящие в других (неаллельных) генах. Это явление называется межгенной супрессией.
Причины возникновения мутаций
По причинам возникновения различают спонтанные и индуцированные мутации.
Спонтанные (самопроизвольные) мутации возникают без видимых причин. Эти мутации иногда рассматривают как ошибки трех Р: процессов репликации, репарации и рекомбинации ДНК. Это означает, что процесс возникновения новых мутаций находится под генетическим контролем организма. Например, известны мутации, которые повышают или понижают частоту других мутаций; следовательно, существуют гены-мутаторы и гены-антимутаторы.
В то же время, частота спонтанных мутаций зависит и от состояния клетки (организма). Например, в условиях стресса частота мутаций может повышаться.
Индуцированные мутации возникают под действием мутагенов.
Мутагены – это разнообразные факторы, которые повышают частоту мутаций.
Впервые индуцированные мутации были получены отечественными генетиками Г.А. Надсоном и Г.С. Филипповым в 1925 г. при облучении дрожжей излучением радия.
Различают несколько классов мутагенов:
– Физические мутагены: ионизирующие излучения, тепловое излучение, ультрафиолетовое излучение.
– Химические мутагены: аналоги азотистых оснований (например, 5-бромурацил), альдегиды, нитриты, метилирующие агенты,гидроксиламин, ионы тяжелых металлов, некоторые лекарственные препараты и средства защиты растений.
– Биологические мутагены: чистая ДНК, вирусы, антивирусные вакцины.
– Аутомутагены – промежуточные продукты обмена веществ (интермедиаты). Например, этиловый спирт сам по себе мутагеном не является. Однако в организме человека он окисляется до ацетальдегида, а это вещество уже является мутагеном.
Общие закономерности мутагенеза
Мутации возникают не мгновенно. Вначале под воздействием мутагенов возникает предмутационное состояние клетки. Различные репарационные системы стремятся устранить это состояние, и тогда мутация не реализуется. Основу репарационных систем составляют различные ферменты, закодированные в генотипе клетки (организма). Таким образом, мутагенез находится под генетическим контролем клетки; это – не физико-химический, а биологический процесс.
Например, ферментные системы репарации вырезают поврежденный участок ДНК, если повреждена только одна нить (эту операцию выполняют ферменты эндонуклеазы), затем вновь достраивается участок ДНК, комплементарный по отношению к сохранившейся нити (эту операцию выполняют ДНК-полимеразы), затем восстановленный участок сшивается с концами нити, оставшимися после вырезания поврежденного участка (эту операцию выполняют лигазы).
Существуют и более тонкие механизмы репарации. Например, при утрате азотистого основания в нуклеотиде происходит его прямое встраивание (это касается аденина и гуанина); метильная группа может просто отщепляться; однонитевые разрывы сшиваются. В некоторых случаях действуют более сложные, малоизученные системы репарации, например, при повреждении обеих нитей ДНК.
Однако при большом числе повреждений ДНК они могут стать необратимыми. Это связано с тем, что: во-первых, репарационные системы могут просто не успевать исправлять повреждения, а во-вторых, могут повреждаться сами ферменты систем репарации, необратимые повреждения ДНК приводят к появлению мутаций – стойких изменений наследственной информации.
Физические мутагены
К физическим мутагенам относятся: ионизирующее излучение (альфа-, бета-, гамма-, нейтронное и рентгеновское излучение), коротковолновое ультрафиолетовое излучение, СВЧ-излучение, действие экстремальных температур.
Действие ионизирующего излучения основано на ионизации компонентов цитоплазмы и ядерного матрикса. При ионизации возникают высокоактивные химические вещества (например, свободные радикалы), которые различным образом действуют на клеточные структуры. Рассмотрим наиболее изученные механизмы мутагенного воздействия ионизирующего излучения.
1. Непосредственное воздействие частиц с высокой энергией на ДНК, которое приводит к ее разрывам: одиночным (под воздействиемгамма-квантов, рентгеновских лучей) или множественных (под воздействием альфа-частиц, нейтронного излучения). Это универсальный механизм возникновения хромосомных перестроек на всех стадиях клеточного цикла, но он действует очень грубо – обычно клетки теряют способность к нормальному делению и погибают. К разрывам ДНК приводит и ультрафиолетовое облучение.
2. Опосредованное воздействие ионизирующих факторов связано с нарушением структуры ферментов, контролирующих репликацию, репарацию и рекомбинацию ДНК. Этот механизм наиболее эффективно действует на синтетической стадии интерфазы. При больших дозах мутагенов клетки погибают. (Поскольку раковые клетки делятся непрерывно, то облучение является универсальным средством подавления развития метастазов при онкологических заболеваниях – непрерывно делящиеся раковые клетки более уязвимы, чем медленно пролиферирующие или непролиферирующие нормальные клетки.)
Опосредованное воздействие ионизирующих факторов индуцирует самые разнообразные генные и хромосомные мутации. При опосредованном действии ионизирующих факторов их мутагенный эффект может быть снижен с помощью специальных веществ – радиопротекторов. К радиопротекторам относятся различные антиоксиданты, взаимодействующие с продуктами ионизации. В то же время, мутагенный эффект может быть усилен, например, высокая температура повышает мутагенный эффект радиации.
3. Особенности мутагенного действия ультрафиолетовых лучей. ДНК интенсивно поглощает жесткий ультрафиолет с длиной волны ≈ 254 нм. Основным продуктом является образование нуклеотидных димеров: два нуклеотида, расположенных рядом в одной цепи ДНК, «замыкаются» сами на себя, образуя пары «тимин–тимин» или «тимин–цитозин». При репликации ДНК напротив такой пары в достраивающейся цепи могут стать два любых нуклеотида, т.е. принцип комплементарности не выполняется. Ультрафиолетовый свет – это сравнительно мягкий мутаген, поэтому его широко используют в селекции растений, облучая проростки.
4. Особенности мутагенного действия экстремальных температур. Собственный мутагенный эффект экстремальных температур не доказан. Однако очень низкие или очень высокие температуры нарушают деление клетки (возникают геномные мутации). Экстремальные температуры усиливают действие других мутагенов, поскольку снижают ферментативную активность репарационных систем.
Химические мутагены
К химическим мутагенам относятся самые разнообразные вещества. Рассмотрим мутагенное действие некоторых из них.
Алкилирующие агенты. Вызывают алкилирование ДНК (например, метилирование, этилирование и т.д.). В результате при репликации ДНК нарушается принцип комплементарности, и происходит замена нуклеотидных пар: ГЦ → АТ; ГЦ → ЦГ; ГЦ → ТА
Некоторые из алкилирующих агентов в природе не встречаются, их не распознают ферменты защитных систем. Такие вещества называютсясупермутагенами (например, N-метил-N-нитрозомочевина). Супермутагены применяются в селекции растений для получения индуцированных мутаций; их используют также как стимуляторы роста (в сверхмалых концентрациях).
Гидроксиламин. Избирательно аминирует цитозин, что также нарушает принцип комплементарности при репликации ДНК. В результате происходит замена ГЦ → АТ.
Нитриты. Осуществляют окислительное дезаминирование гуанина, аденина, цитозина. Также нарушается принцип комплементарности при репликации ДНК. В результате происходит замена АТ → ГЦ.
Аналоги оснований. Это вещества, сходные с «обычными» азотистыми основаниями. Однако они способны образовыватькомплементарные пары с разными «нормальными» основаниями. Например, при репликации ДНК напротив гуанина вместо цитозина достраивается 5-бромурацил (аналог тимина). В дальнейшем напротив 5-бромурацила достраивается аденин, а напротив аденина – обычный тимин. Этот же процесс может идти и в противоположную сторону. В результате происходят замены: ГЦ → АТ или АТ → ГЦ.
Существует множество иных химических факторов, обладающих мутагенным, канцерогенным и тератогенным действием. Например, ионы тяжелых металлов, связываясь с ферментами репликации, репарации и рекомбинации, снижают их ферментативную активность. Таким образом, не являясь собственно мутагенами, ионы тяжелых металлов способствуют появлению мутаций. Кроме того, нужно учесть, что воздействие совершенно разных мутагенов может приводить к сходным результатам.
Опасность загрязнения окружающей среды мутагенами
Проблемы генетической безопасности
Во второй половине ХХ века над биосферой нависла угроза загрязнения мутагенами. Любая популяция способна выдержать лишь определенный груз мутаций. Увеличение частоты мутаций может привести к снижению устойчивости популяций из-за нарушения генетического гомеостаза.
Необходимо дальнейшее усиление эколого-генетического мониторинга – контроля за состоянием окружающей среды на популяционно-генетическом уровне.
В качестве профилактических мер следует использовать развитие «безотходных» технологий, ограничение производства веществ с мутагенным действием, усиление всех видов контроля за состоянием потенциально опасных предприятий: АЭС, химические и микробиологические производства, научно-промышленные установки биотехнологического характера.
Существуют факторы, которые снижают частоту мутаций – антимутагены. К антимутагенам относятся некоторые витамины–антиоксиданты (например, витамин Е, ненасыщенные жирные кислоты), серосодержащие аминокислоты, а также различные биологически активные вещества, которые повышают активность репарационных систем.
Установлено, что мутагены при определенных условиях оказывают канцерогенное и тератогенное действие. Канцерогены – это факторы, провоцирующие развитие онкологических заболеваний; тератогены – это факторы, провоцирующие развитие различных аномалий, уродств. Тератогенный эффект дают многие лекарственные препараты. Например, в 1960-е гг. на Западе широко использовалось снотворноеталидомид, применение которого привело к рождению большого числа детей с недоразвитыми конечностями.
Наряду с тератами – уродствами – часто встречаются морфозы – изменения, которые не ведут к утрате органом его функций. Отличить мутагенное действие от тератогенного сравнительно легко: тераты (уродства) являются ненаследственными модификациями, они предсказуемы (направлены) и не сохраняются в последующих поколениях. Например, серая окраска тела у дрозофилы – это нормальный признак. В то же время известна мутация yellow – желтое тело (эту мутацию легко получить искусственно, обрабатывая родительских особей различными мутагенами; при этом разные мутагены могут давать одинаковый фенотипический эффект). Если же личинкам дрозофилы добавлять в корм азотнокислое серебро, то все эти личинки разовьются в мух с желтым телом. Но, если от этих желтых мух получить потомство и выращивать его на обычной питательной среде, то все потомки вновь станут серыми. Таким образом, в данном случае «пожелтение» тела мух – это не мутация, а модификация, или фенокопия (модификация, по фенотипу копирующая мутацию); азотнокислое серебро в данном случае является не мутагеном, а тератогеном.
Мутон – наименьший участок молекулы ДНК, способный к мутации (равен одному нуклеотиду.)
2).Прогенез.Многофункциональная организация зрелой яйцеклетки. Пространственная упорядоченность цитоплазмы яйца. Значение генома яйцеклетки для начальных стадий онтогенеза.
Прогенез - развитие и созревание половых клеток.
Прогенез включает в себя сперматогенез и овогенез.
Сперматогенез — это развитие и формирование мужских половых клеток. Сперматогенез протекает в извитых канальцах семенников, и его средняя продолжительность от 68 до 75 суток. Сперматогенез у человека начинается с момента полового созревания и продолжается в течении всего активного полового периода в больших количествах.
Стадии сперматогенеза:
размножение;
роста;
созревание-деление;
формирование.
Начальной фазой сперматогенеза является размножениесперматогоний путем митоза, большая часть клеток продолжает делится, а меньшая часть вступает в стадию роста. В этот период клетки растут, накапливают питательные вещества, и потом превращаются в сперматоциты 1-го порядка. Следующая фаза созревание-деление, характеризуется двумя редукционными делениями, без интерфазы. В результате 1-го деления 1сперматоцит 1-го порядка дает начало 2-м сперматоцитам 2-го порядка, а 2-ое деление-созревание приводит к появлению 4 сперматид. Фаза формирования происходит в присутствии тестостерона, происходит преобразование сперматид в сперматозоиды. Ядро сперматиды приобретает видоспецифическую форму, хроматин конденсируется. Комплекс Гольджимигрирует к верхушке головки сперматозоида и образует чехлик и акросому.Центриоли идут к противоположному полюсу, проксимальная центриоль образует колечко в области шейки, а дистальная центриоль дает началоаксонемме — осевой нити сперматозоида. Митохондрии укладываются в промежуточной части хвостика. Микрофиламенты окружают аксонемму в главном отделе хвостика, терминальный отдел хвостика представляет собой ресничку. Акросома содержит сперматолизины (трипсин, гиалуронидаза).
Сперматозоиды — это мелкие, подвижные клетки, размером 30—60 мкм. В сперматозоиде различают головку и хвост. Головка сперматозоида имеет овоидную форму и включает в себя небольшое плотное ядро, окруженное тонким слоем цитоплазмы. Ядра сперматозоидов характеризуются высоким содержанием нуклеопротаминов инуклеогистионов. Передняя половина ядра покрыта плоским мешочком, составляющим "чехлик" сперматозоида. В нем у переднего полюса располагается акросома. Чехлик и акросома являются производными комплекса Гольджи. Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам, способным растворять оболочки, покрывающие яйцеклетку. За головкой имеется кольцевидное сужение. Головка так же, как и хвостовой отдел, покрыта клеточной мембраной.
Хвостовой отдел сперматозоида состоит из связующих, промежуточных, главной и терминальной частей.
В связующей части или шейке располагаются центриоли — проксимальная и дистальная, от которой начинается осевая нить (аксонема). Промежуточная часть содержит 2 центральных и 9 пар периферических микротрубочек, окруженных расположенными по спирали митохондриями. Именно митохондрии обеспечивают энергией двигательную активность сперматозоидов, нарушение которой нередко связано с поражением процесса энергообразования в митохондриях. Главная часть по строению напоминает ресничку. Она окружена тонким фибриллярным влагалищем. Терминальная, или конечная часть содержит единичные сократительные филаменты.
Овогенез — это процесс образования и развития женских половых клеток. Он включает в себя 3 фазы:
размножения;
роста;
созревания.
Фаза размножения начинается в эмбриональном периоде и продолжается в течение 1-го года жизни девочки. К моменту рождения у девочки имеется около 2-х млн клеток. К периоду полового созревания остается около 40 тыс. половых клеток и в последующем 1 раз в 28—32 дня происходит созревание и выход одной яйцеклетки в маточную трубу —овуляция. Овуляция прекращается при наступлении беременности или менопаузы. Сущностью фазы размножения является митотическое деление овогоний.
Фаза роста, в конце 1-го года жизни девочки размножение овогоний останавливается и клетки яичника вступают в фазу малого роста, превращаясь в овоциты 1-го порядка. Наступает 1 блок роста, который снимается с наступлением полового созревания, то есть появлением женских половых гормонов. Далее овоциты 1-го порядка вступают в фазу большого роста.
Фаза созревания, как и во время сперматогенеза, включает в себя два деления, причем второе следует за первым без интеркинеза, что приводит к уменьшению (редукции) числа хромосом вдвое, и набор из становится гаплоидным. При первом делении созревания овоцит 1-го порядка делится, в результате чего образуются овоцит 2-го порядка и небольшое редукционное тельце. Овоцит 2-го порядка получает почти всю массу накопленного желтка и поэтому остается столь же крупным по объему, как и овоцит 1-го порядка. Редукционное же тельце представляет собой мелкую клетку с небольшим количеством цитоплазмы. При втором делении созревания в результате деления овоцита 2-го порядка образуются одна яйцеклетка и второе редукционное тельце. Первое редукционное тельце иногда тоже делится на две одинаковые мелкие клетки. В результате этих преобразований овоцита 1-го порядка образуются одна яйцеклетка и три редукционных тельца.
Яйцеклетки — это наиболее крупные клетки в организме человека, их размер составляет около 130—160 мкм. В цитоплазме яйцеклетки содержатся все органеллы (за исключением клеточного центра) и включения, основной из них — желток (лецитин). В яйцеклетке различают вегетативный полюс, в котором накапливается желток, и анимальный полюс куда смещается ядро. Желток — это включение, которое используется в яйцеклетке в качестве питательного вещества, кроме того под оволеммой содержатся кортикальные гранулы, которые являются производными комплекса Гольджи и образуютоболочку оплодотворения. В ядре яйцеклетки имеется гаплоидный набор хромосом, 22 являются соматическими и 1 (Х) половая. Снаружи яйцеклетка покрыта 3-я оболочками, у человека имеются следующие: оволемма, блестящая оболочка, и оболочка образуемая фолликулярными клетками — "лучистый венец". Блестящая оболочка представляет собой в химическом отношении гликозоаминогликаны и протеогликаны, которые являются продуктом жизнедеятельности яйцеклетки и фолликулярных клеток.
Классификация яйцеклеток:
I. По количеству желтка в цитоплазме:
алецитальныебезжелтковые;
олиголецитальныемаложелтковые;
полилецитальныемногожелтковые.
II. По характеру расположения желтка в цитоплазме:
изолецитальныес равномерным распределением желтка;
центролецитальныежелток располагается в центре яйцеклетки;
телолецитальныежелточные зерна скапливаются у одного полюса яйцеклетки.
Яйцеклетка человека относится к олиголецитальной и изолецитальной.
В ядре яйцеклетки содержатся 23 хромосомы, из них 22 аутосомы и 1 половая Х-хромосома.
В ядре яйцеклетки осуществляется процесс амплифика�ции. Что такое амплификация? Это снятие копий генов РНК с поверхности участков ДНК. Копии генов каких РНК копиру�ются при амплификации? Информационных, транспортных и рибосомных. С этих копий снимаются новые копии. В ко�нечном итоге эти копии свертываются в кольца и выходят из ядра и хранятся до момента оплодотворения. Часто копии ге�нов РНК блокируются белками, называемыми информосомами. Таким образом, в яйцеклетках создается очень мощный трансляционный аппарат.
В цитоплазме яйцеклетки отсутствует клеточный центр, так как он утрачивается в ходе 1-го деления созревания. В то же время хорошо развиты митохондрии, ЭПС. Что касается комплекса ГЬльджи, то он распадается на кортикальные гра�нулы, которые располагаются по периферии яйцеклетки под оволеммой. В этих гранулах содержатся протеолитические ферменты.
Яйцеклетка содержит гиногамоны: гиногамон I и гиногамон II. Гиногамон I — это вещество, которое вызывает поло�жительный хемотаксис у сперматозоидов. Гиногамон II — это вещество, убивающее сперматозоид. В тот момент, когда яй�цеклетка выделяет этот гиногамон, последний соединяется с андрогамоном II, в результате чего сперматозоид обездви�живается и погибает.
В яйцеклетке имеются кальциевые депо. Они предста�вляют собой скопления ионов кальция в цистернах гладкой ЭПС. Желток представлен в яйцеклетке в виде желточных шаров, гранул и желточных пластинок. Желток представля�ет собой питательное вещество, которого яйцеклетке челове�ка хватает на 24 часа автономного существования. Если в те�чение этого времени яйцеклетка не будет оплодотворена, то она погибает.
Яйцеклетка неподвижна. Она передвигается благодаря мускульным сокращениям яйцеводов, мерцаниям ресничек эпителия, выстилающего слизистую оболочку яйцеводов. Ко�личество яйцеклеток также мало по сравнению с количеством сперматозоидов. Так, например, в течение месяца у женщины созревает всего лишь 1 яйцеклетка.
3).Экология споровиков. Морфология, жизненный цикл токсоплазмы. Приобретенный и врожденный токсоплазмоз. Профилактика токсоплазмоза.
K классу споровиков относятся исключительно паразитические простейшие. Паразитируют они в самых различных беспозвоночных и позвоночных животных. Паразитами человека являются споровики из отряда Cосcidiida, подотрядов Eimeridea и Haemosporiidea. B процессе эволюции они приспособились к паразитированию а таких органах как кишечник, печень, почки, кровеносная система, кровь, мышцы и нервная система. Приспособление к паразитизму у споровиков чрезвычайно совершенное. У них отсутствуют специальные органы захвата пищи и питаются они осмотическим путем. Все споровики имеют сложные циклы развития с чередованием бесполой и половой формы размножения, что обеспечивает, с одной стороны, увеличение числа паразитов в данном хозяине, а с другой -образование стадий, служащих для заражения новых особей.
Представитель класса споровиков, отряда кокцидий -Toxoplasma gondii -является паразитом сотен видов млекопитающих и птиц. Он обнаружен во всех континентах и на всех географических широтах. Токсоплазма способна поражать самые разнообразные ткани и клетки своих хозяев. В то же время приобретенный токсоплазмоз может протекать и бессимптомно.
Морфология токсоплазмы
Toxoplasma gondii в стадии внутриклеточного парaзита имеет форму тела в виде дольки апельсина. Один конец токсоплазмы заострен, другой -закруглен. Длина от 4 до 7 мкм. При окраске по Романовскому цитоплaзма голубовато-серая, а расположенное в центре тела ядро -красно-фиолетовое. На переднем конце нахoдится опорноe образование, в стенке которого располагаются спирально закрученные фибриллы -коноид. От него отходят роптрии -трубчатые образования, расширяющиеся на заднем конце. Есть основание полагать, что в роптриях содержатся вещества, обладающие протеолитическими свойствами. Возможно, эти веществa, по другим специальным органоидам -микронемам, изливаются на клеточны е мембраны хозяина. Питание токсоплазмы осуществляется через микропоры - впячивания пелликулы (микроцитостомы). Остальные клеточные структуры типичны (митохондрии, ЭПС и др.). Таким обрaзом, токсоплазма - паразит с очень глубокими и разнообразными приспособлениями.
Цикл развития
|
|
|
 Скачать 2.17 Mb.
Скачать 2.17 Mb.