Биогеография. Биогеография Г. М. Абдурахманов, Д. А. Криволуцкий, Е. Г. Мяло, Г. Н. Огуреева. Серия Высшее образование. М. Академия, 2003. 480 с. Isbn 5769509767 От издателя
 Скачать 13.83 Mb. Скачать 13.83 Mb.
|

|
ГЛАВА IX ОСТРОВНАЯ БИОГЕОГРАФИЯ Начало островной биогеографии было положено замечательным английским натуралистом А.Уоллесом. Работая на островах Малайского архипелага, А.Уоллес поставил перед собой задачу найти границу между Индо-Малайской и Австралийской фаунами и в 1856 г. обнаружил ее между островами Бали и Ломбок; в 1863 г., анализируя фауны птиц на островах, он положил на карту линию разграничения между этими фаунами, получившую название "линия Уоллеса". Правда, в дальнейшем, изучая распределение в том же регионе моллюсков и млекопитающих, М. Вебер провел другую границу (так называемую "линию Вебера"), причем оказалось, что фауна между этими двумя линиями является смешанной (рис. 80). Заслуга А.Уоллеса заключается, однако, не только в проведении разграничительной линии между двумя зоогеографическими областями, но и в объяснении причины ее образования: к западу от "линии Уоллеса" все острова расположены на мелководье континентального шельфа и в историческом прошлом они неоднократно соединялись с материком Азии сухопутными связями. Именно А. Уоллесу биогеография обязана идеей рассматривать не только острова, но и озера, вершины гор, другие изолированные биоты в качестве "островных" экосистем среди окружающих ландшафтов, что было четко высказано им в книге "Островная жизнь" (1880). В настоящее время изучение и анализ островных биот - активно развивающийся раздел биогеографии. Такие биоты характеризуются обилием неизвестных на материках видов, своеобразием флоры и фауны и более простой структурой экосистем. Они удобны для анализа динамики экосистем и часто являются объектами математического моделирования. РАССЕЛЕНИЕ ОБИТАТЕЛЕЙ ОСТРОВОВ Острова Мирового океана различаются по происхождению (океанические и материковые), по размерам и высоте над уровнем моря. Океанические острова никогда не соединялись с сушей, они поднялись со дна океана. К их числу относятся острова коралловые, вулканические и острова складчатых дуг. Материковые острова представляют собой части континентов, отделившиеся от них 349 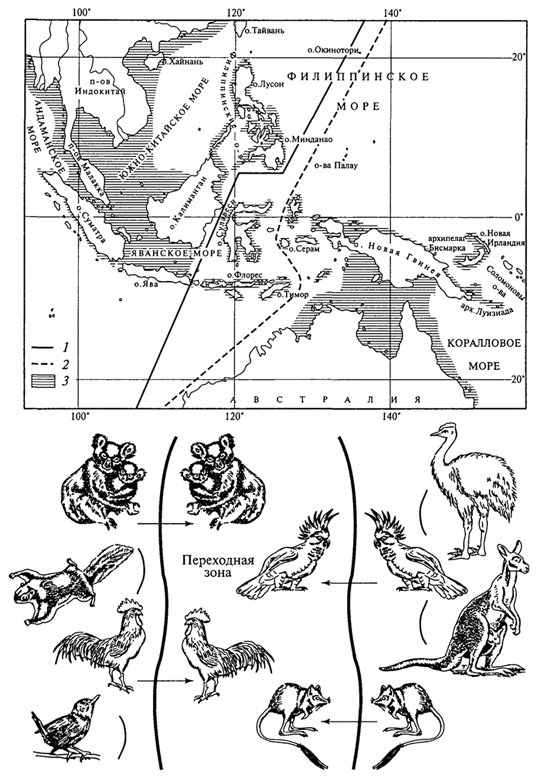 Рис. 80. Распространение некоторых животных в переходной зоне между Индо-Малайской и Австралийской фаунистическими областями (G.Lattin, 1967): 1 - линия Уоллеса; 2 - линия Вебера; 3 -максимальная площадь суши в период последней регрессии моря 350 в ту или иную геологическую эпоху вместе со всей совокупностью живых организмов, которая была свойственна этим участкам континента. В дальнейшем часть первоначального населения вымерла, а на смену ему пришли виды, преодолевшие океан. Дж. Грессит и С. Иошимото (1963) характеризуют способности к распространению животных, относящихся к разным систематическим группам. Они указывают, что из рептилий, например, очень широко распространены сцинки и гекконы, другие же группы ящериц, как и змеи, а также пресноводные рыбы на многих океанических островах отсутствуют. Видимо, представители этих двух групп ящериц хорошо приспособлены к путешествиям на плавающих стволах деревьев или на их скоплениях - "плотах". Из других систематических групп соленую воду очень плохо переносят амфибии. Поэтому возможности их перемещения через океан крайне ограничены. Среди беспозвоночных животных, обитающих на океанических островах, доминируют насекомые, второе по численности место занимают наземные моллюски. Насекомые распространяются всеми названными способами. Виды, распространяемые человеком, заметно отличаются от других, так как на островах они обычно являются синантропными, т.е. используют запасы пищи человека, его одежду (кожееды, моли и др.) и жилища, связаны с домашними животными и сельскохозяйственными растениями. Многие из них имеют космополитические ареалы. Перенос насекомых птицами редок. Соленая вода крайне неблагоприятно действует на них, поэтому виды, преодолевающие океанические пространства вплавь, также немногочисленны. Взрослые растения редко остаются живыми при переносе через океан, хотя плоды некоторых растений могут долго находиться в морской воде, не теряя всхожести, и таким образом расселяться на острова (например, кокосовая пальма, баринтония). Обычно же легкие семена и споры переносятся ветром. Несомненно, важную роль в распространении растений играет человек, разносящий по всему миру, в том числе и по островам, сорняки. Если два острова различной величины недавно отделились от одного и того же континента, то на большем по размерам острове материковая биота в состоянии сохраниться почти полностью, а на меньшем - возможность существования целых таксономических групп, например представителей всего класса млекопитающих, может быть полностью или частично исключена. Ограничивающее влияние размеров острова усиливается по мере уменьшения его территории. Ф.Дарлингтон (1957) для Антильских островов установил еледующее соотношение между размерами острова и числом видов амфибий и рептилий: при уменьшении размеров острова в 10 раз число видов этих групп уменьшается вдвое. 351 Чем ближе остров находится к источнику миграции, тем выше степень насыщения его мигрантами. Этой закономерности подчиняются биоты материковых и океанических островов. Хотя расселение каждой особи носит случайный характер, однако при длительном развитии миграционных процессов они подчиняются закону больших чисел, т.е. статистической вероятности. Ф.Дарлингтон указывал, что если успешно пересекала пространство шириной в 100 миль одна особь из 1000, то при преодолении следующих 100 миль успешно выполнит эту задачу опять одна особь из 1000, миновавших первые 100 миль. Иными словами, достигнет острова, находящегося в 200 милях от источника миграции, уже только одна особь из миллиона, а острова, лежащего в 300 милях от континента, - лишь одна особь из миллиарда. Большое влияние на миграцию оказывает и положение острова по отношению к направлению ветров, которые его пересекают, так называемый ловчий угол для насекомых и семян растений: если остров, вытянутый в длину, расположен перпендикулярно потоку мигрантов, то будет больше шансов, что какой-либо вид его достигнет; если же остров расположен вдоль основного направления движения, то шансов, что организмы, переносимые ветром или морскими течениями, попадут на него, будет значительно меньше. О постепенном вымирании видов на островах свидетельствует тот факт, что небольшие по площади острова континентального происхождения имеют почти чисто океаническую фауну в отличие от крупных островов. Как показали наблюдения Дж. Дайамонда на островах, лежащих у южного побережья Калифорнии, ни на одном из них не было того количества видов, которое могло бы на нем существовать, если бы этот остров являлся частью материка. Через 50 лет после первых наблюдений на этих островах число видов составляло в среднем почти половину возможного. Распространение - лишь первый этап закрепления вида на острове. Он должен пройти полный цикл развития (эцезис) - от появления в данном месте до принесения жизнеспособного потомства. Более толерантные (выносливые) и эвривалентные (обладающие широкими экологическими возможностями) виды легче проходят стадию эцезиса, чем менее толерантные и стенобионтные. Эцезис осуществляется при наличии благоприятных для жизни организма условий: света, тепла, влаги и особенно пищи. Он остается незавершенным чаще всего в результате недоступности необходимых ресурсов. Так, многие насекомые не могут обосноваться на островах из-за отсутствия кормовых растений для их личинок или взрослых особей или в тех случаях, когда нет открытых пресных водоемов, необходимых для прохождения первых стадий развития. Многие птицы не могут выводить птенцов ввиду отсутствия на острове подходящих для гнездования мест. 352 Среди растений наибольшими возможностями для успешного развития и размножения обладают обитатели побережий, семена которых, попадая на остров, встречают там среду, близкую к исходной. Наоборот, обитатели высокогорий имеют небольшие возможности для переноса своих семян через океан (горы часто расположены далеко от берега) и очень мало шансов для прохождения растениями эцезиса на новой родине, где высоких гор может и не быть. Скорость заселения островов невысока. Правда, данные о скорости этого процесса сильно расходятся. Исходя из числа видов цветковых и сосудистых споровых растений во флоре Гавайских островов и геологического возраста этого архипелага Ф. Фос-берг пришел к заключению, что успешное освоение территории растениями осуществлялось здесь в среднем один раз в 20 - 30 тыс. лет, т.е. не чаще, чем 30 - 35 заселений за 1 млн лет. Однако эта скорость не может быть принята для всех островов. Так, низменные атоллы имеют чаще всего возраст 4 - 6 тыс. лет. Если принять скорость заселения, вычисленную Ф.Фосбергом для Гавайских островов, возраст низменных атоллов меньше срока заселения их одним видом. На самом же деле во флоре каждого из таких атоллов насчитывается до нескольких десятков видов, из которых далеко не все были занесены человеком. На поднятый атолл Науру, возраст которого, по определениям геологов, равен примерно 100 тыс. лет, всего 4 - 5 видов растений могли бы проникнуть при скорости, вычисленной Ф.Фосбергом, в то время как флора атолла включает почти 100 видов. Кроме того, следует иметь в виду, что данные о флоре, несомненно, занижены, так как некоторые растения, проникшие на остров, могли впоследствии вымереть. Таким образом, для многих островов скорость их колонизации значительно выше вычисленной Ф. Фосбергом: для низменных атоллов она составляет одно заселение в 200 - 300 лет, для поднятого атолла Науру - примерно одно в 1000 лет. Большая часть биогеографов полагает, что проникновение видов с континента на те или иные острова могло быть облегчено существованием "каменных кладок", т.е. промежуточных островов и островков, которые на протяжении геологической истории то появлялись из вод океана, то вновь скрывались в них. Чаще всего такими временными пристанищами были острова вулканического происхождения. 353 ОСТРОВНЫЕ БИОТЫ Чем остров меньше, тем, как правило, однообразнее на нем жизненные условия. Обеими этими причинами объясняется прямая зависимость, которая наблюдается между размерами острова и числом видов, входящих в состав его биоты. Иллюстрировать это можно на примере гнездящихся птиц (табл. 7). 353 Таблица 7 Число видов гнездящихся птиц на островах различных размеров
Численность обитающих на острове видов зависит и от других причин, в первую очередь от возраста острова и степени его изоляции - удаленности от материка. Необходимым условием видообразования на островах является изоляция. Если непрерывно осуществляется все новый и новый занос особей одного и того же вида на остров, то в результате скрещивания ранее обитавших здесь особей с особями, недавно появившимися, наблюдается некоторая стабилизация особенностей вида, и процесс видообразования резко замедляется. Процесс видообразования связан также с природными особенностями островов. На высоких островах, где на небольшом пространстве наблюдается значительное разнообразие экологических условий, возможности возникновения новых подвидов и видов выше, чем на низких островах при однообразии их природных особенностей. Отсутствие на островах в составе их биот ряда жизненных форм и систематических групп привели к тому, что некоторые виды, попадая на такие острова, проходят так называемую адаптивную радиацию: потомки одного вида, попавшего на остров или архипелаг, сильно изменяются. Так, предок птиц-цветочниц Drepanididae, американский вьюрок, проникнув на Гавайские острова, не встретил здесь конкурентов и дал начало зябликообразным, медососообразным, пищухообразным, дятлообразным и дубоносообразным формам. Возникло несколько родов и множество видов цветочниц, которые заняли разнообразные экологические ниши, что сделало невозможным повторение подобной адаптивной радиации в более поздние времена. Аналогичные примеры адаптивной радиации представляют пальмы на острове Куба, некоторые насекомые и моллюски на Гавайских островах. Примером такой же адаптивной радиации, но не зашедшей столь далеко, служит дерево похутукава (Metrosideros kermadecensis) на острове Рауль (архипелаг Кермадек). В зависимости от условий местообитания оно образует формы, еще не достигшие уровня 354 видовых различий. Так, это приземистый, прижатый к земле кустарник в нижних частях склонов, подвергающихся действию океанического прибоя; невысокий прямостоячий кустарник на вулканической пемзе на днище кальдеры вулкана; прямоствольное дерево в густых насаждениях, гигантское дерево с распростертыми горизонтальными ветвями в разреженных насаждениях, наконец, эпифит и дерево-душитель при поселении на стволах деревьев. В других случаях в результате видообразования на островах возникают монотипные роды и даже семейства. Таковы дерево дегенерия из семейства дегенериевых Degeneriaceae на острове Фиджи, птица кагу на островах Новой Каледонии. Яркая особенность островных биот - большое число эндемиков, часто высокого таксономического ранга. Количество эндемиков и уровень эндемизма зависят от величины островов, их расстояния от континента, разнообразия экологических условий и длительности изоляции. На островах нередко наблюдаются отклонения от обычного облика представителей тех или иных групп: гигантизм или, наоборот, карликовые размеры. Причины этого неясны. Нередко для островов характерны нелетающие птицы и насекомые. Для птиц главную роль в возникновении нелетающих видов играет отсутствие на островах млекопитающих, которые могли бы их истребить; для насекомых - снос ветром и ураганами летающих видов в океан. Чтобы уцелеть, насекомые должны или обладать быстрым полетом, или, наоборот, потерять способность к полету, или затаиваться при ветре в укромных уголках. На многих, даже незначительных по площади, островах имеется немало видов, характеризующихся порхающим, медленным полетом, - златоглазок, комаров, клопов, мелких дневных бабочек, молей. Их обилию способствуют особенности образа жизни, связанного с умением укрываться от порывов ветра. Следовательно, естественный отбор должен был способствовать выживанию нелетающих особей и привести в конце концов к образованию форм, утративших даже органы полета. Наконец, острова способствуют сохранению примитивных (архаичных) форм. Примерами могут служить новозеландская гаттерия, крайне примитивный род насекомоядных - щелезуб с Антильских островов, мадагаскарская хорьковая кошка, или фосса. Объясняется это тем, что в небольших изолированных экосистемах сложившаяся структура сообществ защищена внешними географическими преградами от вторжения новых, более активных групп, победивших в борьбе за существование, которые на материках, успешно расселяясь, вторгаются в уже образованные ранее экосистемы. Отмечено, что изоляция на островах способствует расхождению форм, т.е. географическому видообразованию, но 355 в то же время эволюционный процесс здесь протекает медленнее, чем на материке. Островная биота вообще, кроме эндемизма, отличается бедностью. Объясняется это вымиранием и трудностью проникновения на острова переселенцев. Численность видов подвержена по годам более или менее значительным колебаниям. Но на больших территориях эти подъемы и падения численности могут происходить лишь на какой-то части ареала вида, в то время как в одном месте численность вида падает, в другом она поднимается. Таким образом, если даже все особи известного вида в одной части ареала вымрут, то она сравнительно быстро заселится из смежных частей. На островах же вид легко может исчезнуть полностью. Ясно, что чем меньше территория острова, тем меньше шансов для вида выжить. Изолированность биот отдельных островов - причина их легкого нарушения при изменении человеком природных условий. Вырубка лесов и замена их плантациями как древесных, так и травянистых растений оказываются на островах часто необратимыми, тем более замена лесов полями. Поэтому, приводя примеры вымирания видов под воздействием человека, следует в первую очередь упомянуть обитателей островов: стеллерову (морскую) корову, жившую у побережий Командорских островов, бескрылого чистика (о. Ньюфаундленд), моа (Новая Зеландия) и др. Однако наиболее катастрофическим для фауны и флоры многих островов оказывается завоз человеком (сознательный или бессознательный) новых видов на эти острова. Например, козы на многих островах истребили немало видов растений. Флора о. Св. Елены потеряла из-за коз значительное число видов ранее свойственных ей деревьев; то же отмечено на о-вах Кермадек и др. В настоящее время на многие острова направлены отряды охотников, цель которых резко уменьшить численность этих животных. Растительноядное сумчатое опоссум, завезенное из Австралии в Новую Зеландию, уничтожило леса во многих районах этой страны. Значительный вред фауне островов наносят попавшие туда крысы. Они истребляют яйца и птенцов птиц, гнездящихся на земле. Так, на о. Рауль (архипелаг Кермадек) они полностью истребили кермадекского буревестника, ныне сохранившегося лишь на нескольких небольших по размеру островках, куда крысы не проникли. Для борьбы с крысами, причинявшими значительный ущерб сельскому хозяйству, в частности разведению сахарного тростника и риса, на Кубу и Фиджи был завезен мангуст. Однако, не ограничившись поеданием крыс, этот зверек резко сократил численность птиц, гнездящихся на земле, на Кубе почти уничтожил эндемичный вид щелезуба, а на Фиджи свел к минимуму численность фиджийской игуаны. Огромные опустошения в составе животного населения островов производят свиньи. На Новой Зеландии они истребили представителя 356 монотипичного эндемичного отряда - гаттерию, сохранившуюся лишь на мелких островах, лежащих у побережья Новой Зеландии; почти истребили нелетающих птиц - киви и совиного попугайчика и т.д. Вселение на Новую Зеландию европейского благородного оленя привело к гибели лесов на значительной площади. Оказалось, что истребить этого зверя, завезенного человеком, очень непросто. Не помогли и премии, назначенные за каждое убитое животное. В настоящее время на Новой Зеландии перешли к созданию оленеводческих ферм при жестокой борьбе с оленем, вольно обитающим на острове. Из вышесказанного следует, что любое вселение на остров видов, ранее здесь не существовавших, должно быть тщательно продумано. Необходимо всегда помнить о ранимости природы островов и трудности, а то и полной невозможности ликвидации последствий подобных акций. 357 ЭВОЛЮЦИЯ ОСТРОВНЫХ СООБЩЕСТВ Слово "остров" в широком смысле означает не только участок суши, окруженный водой: озера представляют собой "острова" воды среди суши; вершины гор - "острова" с высокогорными условиями в "океане" более низких территорий; окна в лесном пологе, возникшие при падении деревьев, - "острова" в море древостоя. Можно говорить об "островах" с особым геологическим строением, определенным типом почв или растительности, окруженных иными горными породами, почвами или фитоценозами. Для перечисленных типов "островов" также можно проследить закономерное отношение между видовым богатством и площадью. Существует масса настоящих, реальных островов в океане, множество озер и горных вершин, лесов, окруженных полями, и просто отдельно стоящих деревьев (рис. 81). Даже одна особь или лист представляют собой "острова" для их паразитов. Ранее отмечалось, что существует зависимость числа видов на острове от его площади. Примеры такой зависимости для разных групп организмов приведены на рис. 82. Обычно при графическом ее изображении берут логарифмическую шкалу для обоих параметров, хотя, если соотносить только с логарифмом площади число видов, получаемая линия иногда ближе к прямой. Однако в простых координатах зависимость всегда имеет вид кривой, причем при увеличении площади рост числа видов замедляется. Применяют три подхода к изучению процессов формирования островных биот: а) оценивают разнообразие местообитаний, пригодных на острове для заселения; 357 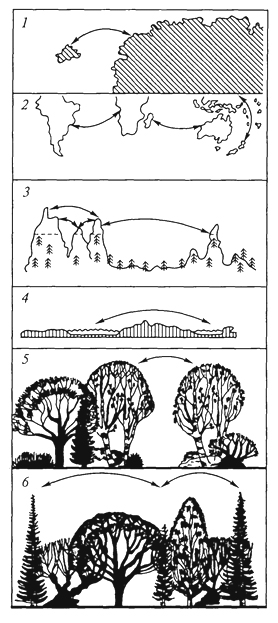 Рис. 81. Острова и "островные" местообитания (П.Мюллер, 1974): 1,2-острова в океане; 3 -"островные" местообитания на вершинах гор; 4- озера как "острова" на суше; 5, <5 - "островные" местообитания для насе- б) обращают внимание на соотношение скорости двух процессов: заселение острова новыми для него видами и вымирание уже поселившихся (теория равновесия); в) выявляют взаимосвязь между заселением острова извне и эволюцией видов на нем самом. В реальной действительности все эти процессы идут одновременно и конкретный результат - своеобразие островной биоты - является итогом, балансом их взаимодействия. М.Бугом с соавторами (1989) проанализировали эти подходы на примере островов различного типа, а также для произвольно выбранных участков материка. Одним из крупных достижений биогеографии нашего времени является создание Р. Мак-Артуром и Е.Уилсоном (1976) "равновесной теории островной биогеографии", основанной на математическом аппарате описания динамичного процесса миграции видов на острова и их вымирания, а также характера устанавливающегося видового равновесия. Суть равновесной теории состоит в том, что количество видов, населяющих остров, определяется равновесием между иммиграцией и вымиранием (рис. 83). Рассмотрим вначале иммиграцию. Представим незаселенный остров. Интенсивность иммиграции видов для него будет высокой, потому что любая поселившаяся здесь особь представляет собой новый для экосистемы вид. По мере увеличения числа обосновавшихся видов скорость внедрения новых (еще не представленных) снижается. Она обратится в нуль, когда на острове поселятся 358 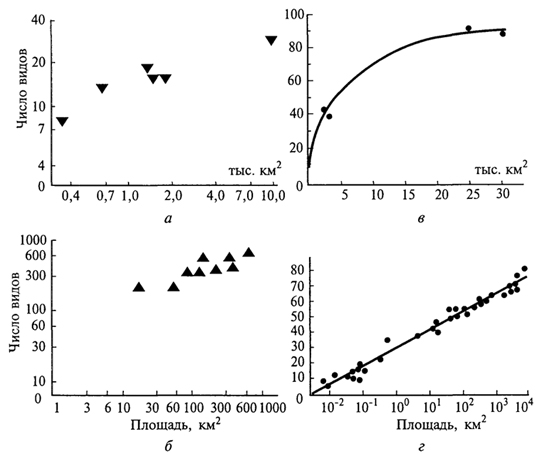 Рис. 82. Графики зависимости "число видов - площадь" для различных групп организмов на островах (R.MacArthur, E.Wilson, 1976): а -местные наземные и водоплавающие пресноводные птицы Гавайских островов; б- сосудистые растения Азорских островов; в -земноводные и пресмыкающиеся островов Вест-Индии; г -птицы Соломоновых островов все виды "исходной совокупности", т.е. все виды соседнего материка или близлежащих островов. Кривая иммиграции зависит от степени удаленности острова от источника его потенциальных заселенцев. Нуль будет отмечаться всегда в одной и той же точке (на острове представлены все виды "исходной совокупности"), но при этом функция в целом будет иметь тем более высокие значения, чем ближе источник иммиграции, т.е. чем больше у мигрантов шансов добраться до острова. Скорость внедрения на крупные острова, как правило, выше, чем на мелкие, так как первые более доступны. Скорость вымирания равна нулю, когда на острове нет ни одного вида, и в целом низка при небольшом их количестве. Однако по мере роста числа обосновавшихся видов она, согласно теории, увеличивается. Чтобы рассмотреть суммарный результат иммиграции и вымирания, наложим оба графика друг на друга. Число видов, соответствующее точке пересечения кривых ,S°, отражает состояние динамического 359 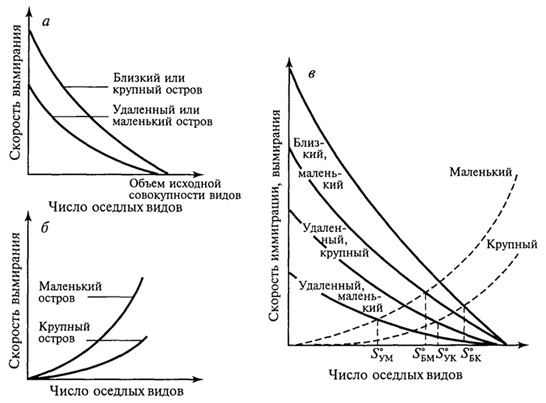 Рис. 83. Равновесная теория островной биогеографии (R.MacArthur, E.Wilson, 1976): а -зависимость скорости иммиграции от числа обосновавшихся видов для крупных и мелких островов, лежащих вблизи и вдали от континента; б -зависимость скорости вымирания от числа обосновавшихся на острове видов для крупных и мелких островов; в -баланс между иммиграцией и вымиранием для крупных и мелких островов и для близких и удаленных от континента; в каждом случае S° обозначает равновесное видовое богатство: S°УМ для удаленных мелких островов, S°БМ -близких мелких островов, S°УМ- удаленных крупных островов, S°БК - близких крупных островов равновесия, т. е. потенциальное видовое богатство острова. Ниже этой точки видовое богатство растет (скорость иммиграции превышает скорость вымирания), а выше - снижается (вымирание интенсивнее иммиграции). Данная теория позволяет высказать ряд предположений о формировании островных сообществ: число видов на острове в конечном итоге должно стабилизироваться; стабилизация числа видов - результат не постоянства видового состава, а непрерывной смены видов, когда одни формы вымирают, а другие - вселяются; на крупных островах больше видов, чем на мелких; видовое богатство острова снижается по мере удаления от источников заселения. 360 Главный вопрос островной биогеографии - существует ли "островной эффект" как таковой или же острова беднее видами просто из-за того, что его территория мала и на ней находится небольшое количество местообитаний? В ряде исследований предпринимались попытки отделить в соотношении "число видов/площадь" для островов компоненты, полностью объясняемые неоднородностью среды обитания, от обусловленных площадью. Например, разнообразие видов рыб в озерах севера штата Висконсин значимо коррелирует как с площадью озера, так и с разнообразием его растительности. Обнаружена также четкая зависимость между видовым богатством птиц и площадью островов у побережья Западной Австралии. Исследование зависимости между числом видов, площадью острова и разнообразием местообитаний для птиц на островах Эгейского моря показало, что разнообразие местообитаний - более важный фактор, чем площадь. Равновесная теория применима и к насекомым-фитофагам (В.Джанзен, 1968): относительно широко распространенные растения можно считать относительно "крупными островами", окруженными "морем" иной флоры. Кроме того, определенный вид растения можно считать "удаленным" от других, если он специфичен по своим морфологическим, биохимическим или прочим биологическим особенностям. Таким образом, из равновесной теории следует, что видовое богатство насекомых будет выше на растениях, имеющих большие ареалы, и ниже на географически изолированных или редких видах, а также на морфологически или биохимически "изолированных" растениях. С равновесной теорией хорошо согласуются данные по островам, составлявшим в прошлом части соединявшегося с материком перешейка, который затем погрузился под воду. Если равновесное число видов в некоторой степени определяется соотношением между скоростью вымирания и площадью острова, следует предполагать, что такие недавно обособившиеся участки суши будут терять виды, пока не установится новое равновесие, соответствующее их размерам. Этот процесс принято называть релаксацией. Так описана зависимость между временем отделения островов Калифорнийского залива от материка и числом встречающихся на них видов ящериц. Внеся поправки на площадь острова, автор продемонстрировал сокращение видового богатства с SOTS видов примерно до 25 за 4000 лет. Равновесная теория предполагает для "острова" не только наличие характерного видового богатства, но и постоянный "круговорот" видов, т.е. непрерывное поселение новых форм и вымирание уже присутствующих, следовательно, состав островной биоты в каждый момент времени должен быть во многом случайным. 361 Давно замечено, что одна из главных особенностей островных биот - дисгармония таксономической и трофической структуры сообщества (Д.Гукер, 1866). Под "дисгармонией" подразумевается иное, чем на материке, относительное участие различных таксонов в составе биоты, в построении трофических сетей островных экосистем. Рассматривая отношения между числом видов и площадью, исследователи уже имели случай убедиться, что организмы, способные хорошо расселяться (такие, как папоротники и птицы) с большей вероятностью заселяют отдельные острова, чем виды, у которых эта способность не столь развита (большинство млекопитающих, практически все хвойные). Потенциальные возможности расселения отдельных видов, конечно, неодинаковы и внутри таких групп. На рис. 84 показаны восточные границы 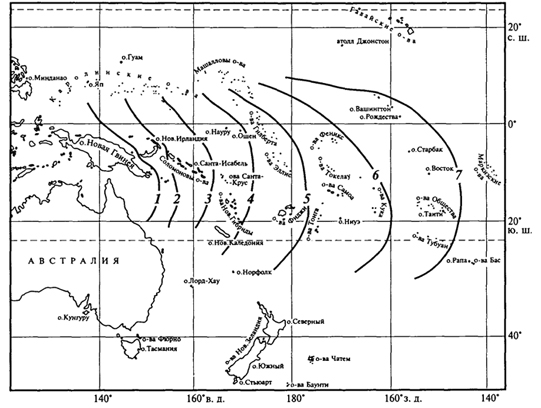 Рис. 84. Восточные границы распространения семейств и подсемейств гнездящихся птиц, встречающихся в Новой Гвинее (J.Davidson, 1945): 1 - не встречаются за пределами Новой Гвинеи (пеликаны, аисты, жаворонки, коньки, райские птицы); 2 - не заходят дальше Новой Британии и архипелага Бисмарка (казуары и фазановые); 3 -не заходят дальше Соломоновых островов (совы, сизоворонки, птицы-носороги, дронго); 4 -не заходят дальше Вануату и Новой Каледонии (поганки, бакланы, скопы, вороны); 5 - не заходят дальше Тонго и Самоа (утки, дрозды, воскоклювые ткачики); 6 -не заходят дальше островов Кука и Общества (сипухи, ласточки и скворцы); к востоку от линии; 7 - встречаются на Маркизских островах и Туамоту цапли, пастушки, голуби, попугаи, кукушки, стрижи, зимородки, славки и мухоловки 362 распространения по островам Тихого океана различных наземных и пресноводных птиц, встречающихся на Новой Гвинее, фауна которых обедняется вдвое на протяжении 2600 км от области расселения. В то же время выяснилось, что большинство видов наземных улиток тихоокеанского региона, будучи очень мелкого размера, легко переносятся с острова на остров, а большинство жуков о. Св. Елены - ксилофаги или подкоровые формы, занесены сюда, вероятнее всего, через море вместе с плавающими стволами деревьев. Однако разные способности к расселению - не единственная причина дисгармонии. Виды с низкой плотностью на единицу площади в естественных условиях всегда будут представлены на островах крайне малочисленными популяциями, для которых вероятность полного исчезновения в результате случайной флуктуации очень высока. На многих островах бросается в глаза отсутствие хищников, популяции которых, как правило, относительно невелики. Например, на архипелаге Тристан-да-Кунья в Южной Атлантике нет ни одного питающегося птицами хищника, не считая тех, что завезены сюда человеком. Хищники могут отсутствовать на островах и по той причине, что их иммиграция в состоянии привести к поселению на острове только в том случае, если там уже обосновалась их добыча (в то время как для видов-жертв подобной зависимости от хищников не существует). Это справедливо также в отношении паразитов, мутуалистов и т.п. Иными словами, дисгармония при заселении островов возникает из-за того, что одни категории организмов более "зависимы", чем другие. 363 ТЕОРИЯ ОСТРОВНОЙ БИОГЕОГРАФИИ И ЗАПОВЕДНОЕ ДЕЛО Теория островной биогеографии, возможно, находит наиболее важное приложение в области охраны природы. Большинство охраняемых территорий и заповедников окружено "океаном" местообитаний, превращенных человеком в непригодные и поэтому "враждебные" для многих видов флоры и фауны. Может ли изучение островов способствовать разработке принципов, применимых при планировании заповедных территорий? Специалисты считают, что да. Наиболее очевидный вывод заключается в невозможности для любой территории как части континуума после превращения ее в изолированный заповедник обеспечить существование стольких видов, сколько обитали в ней до этого. Можно также ожидать, что в крупном заповеднике сохранится больше видов (будет меньшим вымирание), чем в малом. Эти очевидные на первый взгляд 363 выводы требуют некоторых оговорок. Создание изолированного заповедника означает установление границ и, следовательно, появление "пограничных" местообитаний. Значит, в заповеднике смогут обитать виды, которым подходят такие местообитания и которые до выделения заповедника из экологического континуума могли в данной местности отсутствовать. Кроме того, некоторые виды лучше всего чувствуют себя на мелких "островах" и способны сохраниться не в больших, а в малых заповедниках. Поэтому вполне закономерно встает вопрос, создавать ли один крупный заповедник или несколько небольших с такой же суммарной площадью. Если в каждом из малых заповедников обитает один и тот же вид, разумеется, лучше создать один более обширный, тем самым сохраняя больше форм. Однако если регион в целом неоднороден, то в каждом из небольших заповедников будет обитать особый набор видов, и общее количество сохраняемых форм может превзойти таковое в единственном крупном заповеднике. В небольших заповедниках может сохраниться в общей сложности больше видов даже в однородной области, если учесть, что изоляцией может быть достигнута защита от эпидемий, стихийных бедствий, антропогенных воздействий, особенно в окрестностях крупных городов. Для крупных видов хищников, копытных, многих птиц с малой плотностью популяций один крупный заповедник явно предпочтительнее для сохранения вида. При организации заповедников требуется учитывать многие географические особенности среды, в том числе наличие вокруг ландшафтов-аналогов, которые могли бы выполнять функции "буферных зон", "зеленых коридоров" для мигрирующих видов (что было обосновано А.Н.Формозовым в 1939 г.), возможности устойчивого самовозобновления растительного покрова, характер землепользования в округе. Следовательно, основные положения теории островной биогеографии в таких случаях должны быть обязательно учтены. 364 |