Биогеография. Биогеография Г. М. Абдурахманов, Д. А. Криволуцкий, Е. Г. Мяло, Г. Н. Огуреева. Серия Высшее образование. М. Академия, 2003. 480 с. Isbn 5769509767 От издателя
 Скачать 13.83 Mb. Скачать 13.83 Mb.
|

|
ГЛАВА III БИОЦЕНОЗ, БИОГЕОЦЕНОЗ И ЭКОСИСТЕМА Сложная многовидовая система, сформировавшаяся в процессе эволюции живых существ биосферы, получила название биоценоз ("ценоз" - общий), или сообщество. Ельник-зеленомошник, фрагмент красочной луговой степи, своеобразные псаммофильные группировки животных и растений песчаных пустынь - это примеры биоценозов, сочетаний совместно обитающих и взаимодействующих организмов (и их популяций) в различных природных условиях. Различные группы организмов, входящие в состав биоценозов, характеризуются особенностями связей со средой и с другими группами организмов; именно поэтому можно выделять в качестве самостоятельного объекта исследований сообщества микроорганизмов, мелких или крупных млекопитающих, птиц, беспозвоночных, растений. Важнейшая структурная часть биоценоза - растительное сообщество - фитоценоз. Одно из лучших определений фитоценоза принадлежит В. Н.Сукачеву (1954): "Фитоценозом, или растительным сообществом, надо называть всякую совокупность как высших, так и низших растений, обитающих на данном однородном участке земной поверхности, с только им свойственными взаимоотношениями как между собой, так и с условиями местообитания, и поэтому создающую свою особую среду, фитосреду". Фитоценоз в совокупности с животными - это и есть биоценоз. В то же время биоценоз - часть более сложных природных систем, известных как биогеоценозы. Основатель биогеоценологии В.Н.Сукачев рассматривал биогеоценоз как биокосную систему, состоящую из совокупности абиотических условий среды (включая климатические и почвенно-грунтовые) и организмов, образующих биоценоз. В составе биогеоценозов организмы подразделяются на две функциональные трофические группы: автотрофов и гетеротрофов. Автотрофы представлены в основном фототрофами (главным образом растениями), способными на основе использования энергии солнечных лучей создавать органическое вещество, и в меньшей степени - хемотрофами, микроорганизмами, использующими энергию восстановленных химических соединений. Гетеротрофы (животные, бактерии, актиномицеты, грибы, паразитные и сапрофитные растения) получают от автотрофов не только энергетический материал, но и необходимые для 78 жизни вещества. Гетеротрофы, особенно сапротрофы, осуществляют минерализацию отмерших органов растений, а некоторые из них - фиксацию атмосферного азота. Гетеротрофы в значительной степени обеспечивают автотрофов необходимыми для них элементами минерального питания и углекислым газом, а также участвуют в трансформации энергии и биологическом круговороте веществ. Часто наряду с термином "биогеоценоз" используют термин "экосистема". Экосистема и биогеоценоз по существу - близкие понятия. Однако экосистема понимается как безразмерное образование. Как экосистему, например, рассматривают гниющий пень в лесу, отдельные деревья с живущими на нем организмами, лесной фитоценоз, в котором эти деревья и пень расположены, лесной массив, в который входит ряд фитоценозов, лесную зону и т.д. Экосистемой может быть не только биогеоценоз, но и зависимые от биогеоценозов природные биокосные системы, в которых организмы представлены лишь гетеротрофами, а также такие созданные человеком биокосные системы, как зернохранилище, аквариум и др. Биогеоценоз же всегда понимают как хорологическую пространственную единицу, имеющую определенные границы, очерченные входящими в его состав фитоценозами. "Биогеоценоз - это экосистема в границах фитоценоза" (Е. М.Лавренко, Н.В.Дылис, 1968). Изучение биоценоза требует выявления закономерностей его организации. В.Н.Сукачев (1975), характеризуя закономерности организации фитоценозов, отмечал, что основным свойством всякого сообщества является многостороннее взаимное влияние растений друг на друга. Отношения между членами сообщества зависят от экологических свойств растений, входящих в состав сообществ, и от условий среды. Эволюция растений протекала, с одной стороны, под влиянием воздействия условий среды и этих взаимоотношений - с другой. Каждый вид формировался в соответствии с условиями сообщества, одновременно с выработкой тех или иных экологических особенностей растений; как логическое следствие этого шла выработка и взаимоотношений между видами в сообществе. Все это в равной мере относится и ко всем другим группам организмов. Именно связи и взаимодействия - фундаментальная черта биоценозов, они определяют их целостность. В организации биоценозов выделяют трофические, средообразующие, конкурентные и некоторые другие связи. Наиболее значимы конкурентные взаимоотношения, видимо, наиболее всеохватывающие в природе, поскольку ресурсы (средства для жизни) всегда ограничены (свет, влага, элементы минерального питания для растений, пища, в меньшей степени укрытия, территория - для животных). Сложные механизмы конкурентных взаимоотношений 79 в биоценозах - предмет детальных исследований. Среди конкурентных взаимоотношений различают внутривидовые и межвидовые. Внутривидовая конкуренция - конкуренция между особями одной популяции, имеющими потребности в одних и тех же ресурсах для их роста, размножения и выживания. Внутривидовая конкуренция влияет преимущественно на плодовитость и смертность, регулируя таким образом численность популяции. Так, на любой лесосеке, на одном гектаре насчитываются сотни тысяч экземпляров всходов или сеянцев какой-либо древесной породы, однако стволов во взрослом состоянии на той же площади остается всего лишь несколько сотен. Высокая смертность всходов и молодых деревьев на ранних этапах зарастания лесосек и формирования искусственных лесных насаждений давно изучена лесоводами, а по отношению к травянистым растениям - и агрономами. Существует известная зависимость между скоростью роста древесной породы, возрастом древостоя и выпадением стволов. Число погибших деревьев возрастает с момента смыкания крон и, достигнув максимума в определенном возрасте древостоя, сходит на нет (рис. 7). Ход изреживания стволов с возрастом на единицу площади протекает по-разному у светолюбивых и тенелюбивых пород: первые характеризуются более высокой убылью по мере роста. Один из парадоксальных результатов внутривидовой конкуренции у растений - уменьшение плотности популяции вида в более благоприятных условиях, при лучшей обеспеченности ресурсами. Казалось бы, в таких условиях интенсивность убыли должна быть снижена. Однако за счет более быстрого роста и достижения больших индивидуальных размеров число взрослых стволов на единицу 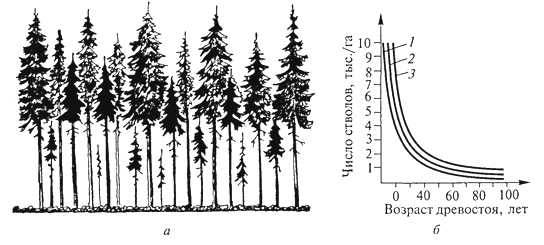 Рис. 7. Самоизреживание в древесных насаждениях: а- господствующие и угнетенные деревья в ельнике; б- ход изреживания стволов с возрастом у сосны (1), березы (2) и ели (3) (Г.Ф.Морозов, 1928) 80 площади ниже, а смертность выше. Следовательно, внутривидовая конкуренция ограничивает численность популяции вида, которая, в свою очередь, варьируется в разных местообитаниях в зависимости от обеспеченности ресурсами. Одним из наглядных выражений внутривидовой конкуренции является дифференциация одновозрастного древостоя по высоте, диаметру ствола, мощности крон. Известная классификация Крафта делит все деревья на пять классов. Если к классу I относятся господствующие деревья, особенно мощные кроны которых расположены над общим древесным пологом и исключительно хорошо развиты, то к классу II - деревья, составляющие основу лесного полога, кроны которых развиты несколько хуже, чем у деревьев I класса, а к V классу относятся деревья, кроны которых полностью размещаются ниже общего полога, это угнетенные, отмирающие или уже отмершие деревья (В.Н.Сукачев, 1928). Между классами разных деревьев существуют постепенные переходы. Внутривидовая конкуренция сказывается на плодоношении деревьев. Если принять плодоношение у ели для деревьев I класса за 100%, то плодоношение для остальных классов выразится в следующих цифрах (в %): II класс - 88, III - 37, IV - 0,5, V - 0, т.е. деревья не плодоносят. Таким образом, как писал Г.Ф. Морозов (1925), "оставляют после себя потомство только победившие в борьбе за существование, только наделенные наибольшей индивидуальной силой роста". В любом биоценозе выражены и межвидовые конкурентные отношения. Возможность совместного сосуществования многих видов в одном биоценозе определяется их эколого-биологической индивидуальностью и дифференциацией условий существования в пространстве (почва, подстилка, осветленные "окна" и пятна под сомкнутыми кронами в лесу, различия в освещенности от верхнего полога растительности до припочвенных слоев) и во времени (хотя бы от сезона к сезону). Длительное существование в составе многовидового сообщества привело в эволюционном плане к тому, что каждый вид пространственно и функционально занимает определенное положение в составе биоценоза, или особую, свойственную ему экологическую нишу. Представление об экологических нишах введено зоологами. В опытах Г. Ф.Гаузе с простейшими животными (Paramecium caudatum, P. aurelid) при совместном существовании двух видов в однородной среде численность одного возрастала, а второй вид исчезал (рис. 8). Это послужило поводом для обоснования принципа конкурентного исключения одного вида другим: два вида на одной и той же территории не в состоянии занимать длительное время одну и ту же экологическую нишу. Первоначально экологическая ниша трактовалась только как ниша пространственная, т.е. как место видовой популяции в биоценозе. Позже экологическую 81 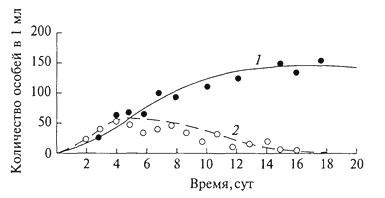 Рис. 8. Динамика конкурентных отношений двух видов инфузорий Рагатесшт aurelia (1) и Р. caudatum (2) в экспериментальной культуре (Г. Ф. Гаузе, 1934) нишу стали рассматривать как интегрированное выражение места вида в биоценозе, в том числе по отношению к факторам внешней среды, и его функциональную роль. По образному выражению американского эколога Ю. Одума (1975), "экологическая ниша некоторого организма зависит не только от того, где он живет, но и от того, что он делает (как он преобразует энергию, каково его поведение, как он реагирует на физическую и биологическую среду) и как он ограничен другими видами". Следовательно, в сложившихся биоценозах существует столько экологических ниш, сколько видов входит в их состав. Конкурентные взаимоотношения между видами смягчены благодаря процессам сопряженной эволюции и 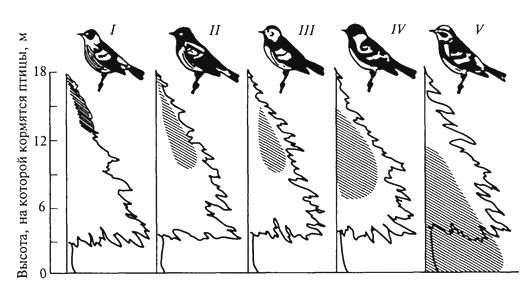 Рис. 9. Локализация мест кормежки пяти видов американских славок: (Dendroica) в еловых лесах штата Мэн (R. H. MacArthur, 1958) I -D. tignna, II - D. fusca, III- D. virens, IV - D. castanea, V- D. coronata 82 взаимной адаптации. Данное положение находится в соответствии с представлением об экологической индивидуальности видов и является одной из важных предпосылок, определяющих возможность их совместного существования. В то же время четкого разграничения экологических ниш в биоценозах нет и не может быть, они всегда частично перекрываются (рис. 9). Межвидовую конкуренцию рассматривают как движущую силу эволюции в направлении выживания разных видов в одном биоценозе путем дифференциации взаимоотношений видов со средой и друг с другом. Таким образом, биоценоз - "это система взаимодействующих, дифференцированных по экологическим нишам, частично конкурирующих друг с другом видов. Виды, входящие в состав одного и того же сообщества, эволюционировали в направлении дифференциации их экологических ниш" (Р.Уиттекер, 1980). 83 ЭКОТОП, БИОТОП, МЕСТООБИТАНИЕ Совокупность факторов окружающей среды, влияющих на состав и структуру биоценозов, носит название местообитание, экотоп или биотоп. Комплексные факторы среды могут быть подразделены на элементарные факторы. По отношению к действию каждого из них у организмов существуют различные адаптации, в связи с чем виды могут быть объединены в экологические группы. Так, по отношению к свету растения подразделяются на светолюбивые (или гелиофиты), тенелюбивые (сциофиты) и теневыносливые (переносят затенение, но хорошо растут на свету); по отношению к влаге - на гидрофиты - водные и наземно-водные, частично погруженные в воду растения мелководий; гигрофиты - наземные растения, связанные с повышенной влажностью воздуха и часто влажными почвами; мезофиты - способные переносить краткую и не очень сильную засуху; ксерофиты - растения местообитаний с недостаточным атмосферным и почвенным увлажнением. По отношению к температуре все разнообразие организмов подразделяется на пойкилотермные (жизнедеятельность которых зависит от тепла, поступающего извне, а температура определяется внешней средой) и гомойотермные, способные поддерживать постоянную внутреннюю температуру тела. К пойкилотермным относятся все микроорганизмы, растения, беспозвоночные животные, к гомойотермным - птицы и млекопитающие. Не менее значимы эдафические факторы: химические, физические и механические особенности почв, играющие огромную роль в распределении растений и животных, в том числе почвенных микроорганизмов. Среди микро- и макроэлементов, поглощаемых корнями растений из почвы, особенно большую роль играют 83 фосфор, калий и азот. Дефицит в почве доступных форм азота - один из важнейших факторов, оказывающих влияние на состояние растений, их конкурентоспособность. Высока чувствительность различных видов растений к рН почвенных растворов; давно известны различия в составе и структуре биоценозов, формирующихся на известняках и силикатных породах. Выделяют экологические группы ацидофильных и кальцефильных растений. К ацидофильным растениям относятся, например, сфагновые мхи, основные ценозообразователи таежных и тундровых болот, кальцефилы - бук лесной, ясень и др. Существенно различаются местообитания на песчаных и суглинистых или глинистых почвах. Песок - подвижный субстрат, имеющий рыхлую структуру, в связи с чем обитатели песков, растения и животные, обладают специфическими особенностями, позволяющими им осваивать этот субстрат. Растения способны давать придаточные корни при засыпании песком нижних частей их стеблей, у многих из них имеются длинные корневища, скрепляющие субстрат, часто корни одеты чехликами из частиц песка, надежно защищающими их от высыхания. Плоды покрыты пленчатой оболочкой, образующей пузырь, или представляют собой шарики с упругими волосками на поверхности, что дает им возможность перекатываться по песку. Это особая экологическая группа - псаммофиты. Животные, обитающие в песках, способны быстро в них зарываться. Ящерицы-круглоголовки делают это с такой быстротой и так хорошо и быстро заравнивают песок, что, даже наблюдая за этим процессом, трудно обнаружить место укрытия. На лапках многих животных, обитающих в песках, имеются щеточки из волосков или роговые чешуйки, увеличивающие поверхность опоры. Это приспособление для более успешного рытья нор в рыхлом грунте, а также, видимо, для быстрого передвижения по песку. К эдафическим факторам относят и снежный покров, который, укрывая поверхность почвы, способствует сохранению тепла в ней. В многоснежных районах возможно подснежное развитие растений, которые после этого зацветают ранней весной. Животных по отношению к снежному покрову А.Н.Формозов предложил разделить на две группы: хионофобов - "снегоненавистников", для которых снежный покров неблагоприятен, так как они ведут надснежный образ жизни, и хионофилов - "снеголюбов", которые обитают под снегом, снежный покров создает для них более благоприятные условия существования. Орографические факторы, за редким исключением, прямо не воздействуют на организмы, но существенно влияют на перераспределение тепла и влаги. Так, значительны различия в условиях местообитания на склонах северной и южной экспозиции. В.В.Алехин и Г.Вальтер сформулировали правило предварения, 84 согласно которому виды южного распространения на севере тяготеют к склонам южной экспозиции, а виды северного распространения, наоборот, на юге предпочитают склоны северной экспозиции. Огромное воздействие на формирование условий местообитания и их трансформацию оказывает деятельность человека. Различные формы и виды воздействия (вырубки, распашки, выпас домашнего скота, разработка месторождений полезных ископаемых, прокладка дорог и т.д.) всегда в той или иной степени изменяют условия местообитания. Глубокой трансформации подвергаются местообитания при добыче полезных ископаемых, особенно в тех случаях, когда на поверхность выводятся коренные породы, при загрязнении поверхности продуктами нефтедобычи. Экологические условия местообитания складываются за счет совокупного действия ряда факторов; особенно большое значение в формировании экологического режима имеют влажность и температура. Иногда различают факторы прямого действия (свет, тепло, влага и др.) и косвенного действия (почвообразующие породы, рельеф). Можно вслед за Г.Н.Высоцким выделить собственно экологические условия (влага, свет, тепло, обеспеченность элементами минерального питания для растений, пища, наличие убежищ для животных) и топологические (почвообразующие породы, уровень грунтовых вод и т.п.) условия. Конкурентно более сильные виды осваивают в ряду прочих более теплые и влажные местообитания, лучше обеспеченные пищей, участки с наиболее благоприятными экологическими условиями (рис. 10). Каждый биоценоз воздействует на среду обитания, существенным образом изменяя температурный режим, влажность воздуха, количество света, почвенные условия и т.д. Огромная роль во влиянии биоценозов на условия среды принадлежит растениям, формирующим свою фитосреду, хотя и вклад животного населения значителен. Надземные органы растений, образующие более или менее сомкнутые покровы, частично перехватывают поступающие в биоценоз солнечную радиацию и атмосферные осадки, изменяют световой и тепловой режимы, а также обеспеченность водой. Вода, проникая в почву в период активной вегетации растений, в большинстве биоценозов поглощается их корнями и расходуется на транспирацию, а также на построение новых органов. Расход воды может быть значительным, например одно дерево березы в солнечный день испаряет 100 л воды, а бука за вегетационный период - 9000 л (В. Лархер, 1978). Благодаря этому растения оказывают воздействие на уровень почвенно-грунтовых вод. Его понижение из-за интенсивной транспирации растений нередко приостанавливает болотообразовательный процесс, что дает возможность поселяться мезофильным растениям с неглубокой 85 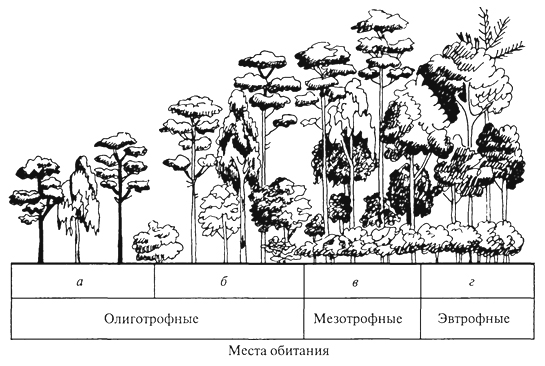 Рис. 10. Взаимосвязь типов леса и почвенных условий западнее Днепра: а -бор на песчаной почве (сосна с единичной березой); б -суборь на супеси (нижний древесный ярус образован дубом); в -сугрудок на суглинке (нижний ярус древостоя образован грабом); г -груд на серых лесных почвах, подстилаемых лëссом (верхний древесный ярус образован дубом, нижний - грабом; развит ярус кустарников) (П.С. Погребняк, 1968) корневой системой. Некоторые виды, обладающие сильно развитой воздухоносной тканью, например тростник, способны аэрировать почву, насыщенную водой (К. А. Куркин, 1976). Внутри биоценоза снижаются сила ветра и интенсивность газо- и теплообмена. Большое значение в образовании фитосреды имеет ежегодное поступление в почву и на ее поверхность отмерших органов растений, которые могут накапливаться в виде подстилки, торфа, гумуса. Продукты разложения отмерших организмов играют исключительно важную роль в определении направления и интенсивности почвообразовательного процесса. Формирование фитосреды наиболее резко выражено в лесах, особенно с хорошо развитым сомкнутым древостоем, где на поверхность почвы ежегодно поступает значительное количество отмерших листьев (опада), образующих особый, очень важный для произрастания растений и обитания многих видов животных горизонт - подстилку. Примером различия в интегральном воздействии отдельных видов на среду и, как следствие этого, на другие компоненты биогеоценоза могут быть результаты наблюдений над посадками 86 ели и сосны в Подмосковье (Н. В. Дылис и др., 1973). Здесь в одинаковых почвенных условиях было посажено равное число на единицу площади молодых деревьев ели и сосны. В возрасте 30 лет культура ели представляла собой чистое насаждение с очень плотным и равномерно сомкнутым пологом густоохвоенных крон ели, совершенно лишенное других растений: кустарников, трав, мхов. В культуре сосны того же возраста древесный полог был также хорошо сомкнут, но пропускал достаточно света для того, чтобы под сосной смогли произрастать 8 видов кустарников (преобладала малина) и 36 видов трав. За 30 лет в сосновом насаждении отмерло 3/4 посаженных деревьев, а в еловом - только половина. Большие различия между посадками наблюдались также в составе других биокомпонентов (животных, микроорганизмов). Различия в воздействии ели и сосны на среду хорошо заметны и в более старых культурах. Сосняк отличался от ельника не только сомкнутостью крон, но и их сквозистостью. Под полог сосны при высоте солнца 45° поступало 40 % прямой солнечной радиации, под полог ели - 30 %, в утренние и вечерние часы под полог ели проникало в 4 -5 раз меньше солнечной радиации, чем под полог сосны. В связи с этим в сосняке лучше развит кустарниковый и травяной покров, но менее выражен почвенный покров из мхов. Оподзоленность почвы в сосняках выражена меньше. Масса подстилки в ельнике больше (8,7 т/га), чем в сосняках (4,2 т/га), что связано с медленным разложением опада ели. Очень большие различия установлены в составе зоокомпонентов: дождевых червей на 1 м2 было в ельнике 131, в сосняке - 221. Велико и средообразующее воздействие животных, особенно различных фитофагов и землероев. Воздействие животных-фитофагов заключается не только в отчуждении и частичной переработке растительной массы, оно оказывает ограничивающее влияние на рост и развитие поедаемых растений, способствуя таким образом уменьшению средообразующей роли одних и увеличению роли других видов. Пример влияния животных на лесные биоценозы - деятельность насекомых-фитофагов. Листо- и хвоегрызущие насекомые нападают в разные годы с различной интенсивностью. У деревьев, ослабленных неблагоприятными условиями, например засухой, меньше сопротивляемость к повреждениям листогрызущих насекомых. В результате массовых вспышек размножения насекомых нередко происходит полная дефолиация деревьев. Так, по данным Р. И.Злотина и К.С.Ходашевой (1974), дубовая листовертка в лесостепных дубравах Курской области вызывала дефолиацию на значительной площади лесов в 1964, 1966- 1968 гг. Для лиственных пород достоверно неизвестна массовая гибель деревьев в результате нападения листогрызущих форм, хвойные же породы нередко гибнут на больших площадях. При этом не исключено, 87 что массовая гибель свойственна деревьям в большей степени в искусственных сообществах или в сообществах, возобновляющихся на вырубках и пожарищах. Средообразующее воздействие животных, в первую очередь млекопитающих и саранчовых, ярко проявляется в степях, где ими нередко травостой уничтожается полностью. Стадные саранчовые до тех пор, пока численность их не была подавлена человеком, появлялись в разных районах степей гигантскими стаями. Как грызуны, так и саранчовые - полифаги, в годы массовых размножений они широко используют и те растения, которые в годы пониженной численности относятся к числу второстепенных кормов. При отсутствии выпаса в степях накапливается влагоемкая подстилка, что приводит к гибели компонентов степи и к ее забурьяниванию, на что указывал еще И. К. Пачоский. Поэтому умеренный выпас копытных необходим для самого существования степей. Многие виды млекопитающих связаны в своей жизнедеятельности с почвой как средой обитания; роющая деятельность животных оказывает прямое и косвенное воздействие на экотоп. Нарушая корневые системы растений при рытье убежищ и ходов, землерои изменяют взаимоотношения между видами, способствуя развитию определенных жизненных форм и видов растений. Перерывая почву, животные выносят ее более глубокие слои на поверхность, происходит перемешивание с верхними слоями, улучшаются условия аэрации; в норах и на участках кормления происходит обогащение почвы азотом. Кроме того, многие землерои формируют специфический мелко- и микробугорковый нано- и микрорельеф. Все это вызывает дифференциацию экологических условий в пространстве. Наиболее существен вклад землероев в формирование условий местообитания в лугах, степях и прериях. 88 |