Биогеография. Биогеография Г. М. Абдурахманов, Д. А. Криволуцкий, Е. Г. Мяло, Г. Н. Огуреева. Серия Высшее образование. М. Академия, 2003. 480 с. Isbn 5769509767 От издателя
 Скачать 13.83 Mb. Скачать 13.83 Mb.
|

|
СУКЦЕССИИ Происходящие во времени смены одного биоценоза другим получили название сукцессии. Одно из важнейших свойств биоценоза - способность реагировать на направленное изменение условий существования последовательными сменами. Так, под воздействием непрерывного, медленного, но неуклонного повышения уровня грунтовых вод на какой-то территории происходят изменения влажности почв, миграции солей, условий аэрации, что существенным образом сказывается на корнеобитаемом слое. В связи с этим происходит изменение ценотически сложившихся отношений 108 в биоценозе, численность одних видов снижается вплоть до их исчезновения, происходит постепенное внедрение новых видов извне, с течением времени возникает новый биоценоз. Направленная и непрерывная последовательность появления и исчезновения популяций разных видов, изменение видового состава биоценоза во времени, приводящая к последовательной смене одного биоценоза другим, и есть сукцессия, а отдельные биоценозы, соответствующие разным этапам развития биоценозов, - это стадии сукцессии. Глубокая разработка теории сукцессии связана с именами американских ученых - Г. Коулса и Ф. Клементса; большой вклад в развитие этой теории внесли крупнейшие отечественные ученые - Г.Ф. Морозов, И. К. Пачоский, Г. Н. Высоцкий и особенно В. Н. Сукачев. Наиболее полно изучены сукцессии растительности, что объясняется огромной ролью растений в формировании структуры биоценоза и развитии сукцессии, их способностью в известной степени направлять ее ход, контролируя возможности существования для других организмов. Это не исключает роли животных, особенно фитофагов и землероев, в развитии тех или иных стадий сукцессии, их воздействия на численность и существование тех или иных видов. Сукцессии разномасштабны как по площади, которую они охватывают, так и по продолжительности. Это смены, например, при зарастании временных водоемов, склонов молодых оврагов, гарей, залежей, сукцессии на выбросах земли из нор кротов, сусликов. Длительные вековые смены связаны с крупномасштабными климатическими изменениями. Принципиально важно разделение сукцессии на первичные и вторичные. Первичные сукцессии развиваются на субстратах, где ранее жизнь отсутствовала, это сопряженные процессы почвообразования и постепенного формирования растительности и животного населения, очень медленные, протекающие столетиями. Таковы смены растительности на породах, обнажившихся после таяния ледников или на побережьях вследствие отступания моря. Так, на обнажившихся скальных породах развитие биоценоза начинается с поселения накипных лишайников или пионерных видов в трещинах скал. Это, как правило, очень длительная стадия, для которой характерно также присутствие водорослей, некоторых простейших, насекомых и клещей. По мере разрушения твердой породы и постепенного образования тонкого рыхлого слоя поселяются другие группы лишайников, различные виды мхов, увеличивается разнообразие животных, продолжается формирование почвы, с развитием которой происходит вселение сосудистых растений, идет обогащение животного населения. Таким образом, в ходе первичной сукцессии идет процесс первоначального становления биоценоза, связанный с заселением 109 нового пространства пионерными формами жизни, отбором видов в процессе освоения конкретных условий среды и конкуренцией за средства жизни - эцезис. Рассмотрим развитие первичной сукцессии, например, на молодых ледниковых отложениях в Канаде (М.Бигон, Д.Харпер, К.Таундсен, 1989). Необыкновенно быстрое отступление ледника отмечено на юго-востоке Аляски, в районе бухты Глейшер. Примерно с 1750 г. ледник отошел здесь почти на 100 км, оставив морены, возраст которых может быть оценен по числу годичных колец наиболее старых поселившихся на них деревьев. На последней моренной гряде максимальный возраст деревьев примерно 200 лет, а по мере приближения к леднику снижается. Первыми на молодых ледниковых отложениях поселяются мхи и отдельные сосудистые растения с поверхностной корневой системой, в частности куропаточья трава, затем - ивы, сначала стелющиеся, позже - кустарниковые виды. Вскоре в сукцессию вовлекается ольха, и примерно через 50 лет возникают ее плотные заросли высотой до 10 м, в которых единично встречается тополь. Ольха вытесняется ситхинской елью и в конечном итоге образуется густой смешанный лес с видами тсуги. Важная движущая сила сукцессии в данном случае - изменение почвы первыми поселенцами. И куропаточья трава, и ольха имеют симбионтов, фиксирующих атмосферный азот, что приводит к накоплению в почве его запасов. Ольха также способствует сильному подкислению почвы, снижая за 50 лет рН в ее верхних горизонтах приблизительно с 8,0 до 5,0. После этого ситхинская ель, используя накопленный азот, может поселяться и замещать ольху. Постепенное накопление в почве углерода ведет к развитию мелкокомковатой структуры, повышению аэрации и влагоëмкости субстрата. На хуже дренированных склонах сукцессия не всегда заканчивается елово-тсуговым лесом. Во влажных местах, с худшим дренажем, бурно развиваются сфагновые мхи, накапливающие воду и сильно подкисляющие почву, которая испытывает дефицит кислорода; при этом большинство деревьев гибнет. Единственный вид, способный выдержать плохую аэрацию субстрата возникающего болота, - сосна скрученная. Таким образом, в развитии сукцессии важное значение приобретают экологические различия; формирование растительности на поздних стадиях сукцессии контролируется почвенно-геоморфологическими условиями. Вторичные сукцессии - это смены биоценозов, начинающиеся от сформированной растительности. Там, где есть почвы с запасом семян, спор, различных вегетативных зачатков, развиваются вторичные сукцессии, как правило, протекающие намного быстрее первичных. Вторичные сукцессии развиваются под воздействием внешних по отношению к биоценозу факторов (изменение 110 уровня грунтовых вод, подтопление, накопление токсичных солей в верхних почвенных горизонтах, выпас, вырубки, пожары) и в результате деятельности биоты. Если внешнее воздействие вызывает полное или почти полное уничтожение растительности, то вторичная сукцессия развивается в направлении восстановления нарушенного (уничтоженного) биоценоза. Так, на месте пожара или вырубки начинается восстановительная сукцессия, смена биоценозов идет в направлении к состоянию, близкому к исходному, существовавшему до нарушения. Этот процесс восстановления в ходе сукцессии исходных биоценозов получил название демутации. В ряде случаев, например под воздействием выпаса, происходит постепенная смена биоценозов, сопровождающаяся упрощением структуры, обеднением видового состава и на крайних стадиях характеризующаяся почти полным исчезновением растительности (сильно выбитые пастбища). Этот процесс известен под названием дигрессия. Многообразие сукцессии потребовало их дифференциации. В зависимости от того, обусловлена ли сукцессия изменением условий среды под воздействием жизнедеятельности самих организмов или определяется внешними по отношению к биоценозу причинами, их подразделяют на автогенетические (эндоэкогенетические, по В.Н.Сукачеву) и аллогенные (экзоэкогенетические). В основе эндоэкогенетических сукцессии лежат изменения связей в биогеоценозах, вызывающие их саморазвитие, экзоэкогенетические сукцессии обусловлены внешними, абиотическими причинами. Яркий пример автогенетических смен - сукцессии при зарастании озер. Не менее интересны смены при развитии сфагновых болот, формирующихся на месте низинных (рис. 17). Отложение торфа в низинных болотах обусловливает в конце концов повышение его поверхности и отрыв от притока грунтовых вод, что вызывает обеднение верхней части торфяной толщи элементами минерального питания. В этих условиях происходит разрастание сфагновых мхов, мощных эдификаторов, формирующих сфагновые ковры с очень бедным видовым составом. На этой стадии торфяная масса, образуемая растительными остатками, постепенно поднимает и уплотняет жидкий горизонт и создает условия, благоприятствующие разрастанию многолетнего травянистого растения - пушицы. Для этой стадии характерен гораздо более устойчивый моховой покров, лежащий на уплотненном, во всей толще спрессовавшемся торфе, образованном другими видами сфагнума. Дальнейшее накопление торфа ведет к созданию условий, благоприятных для произрастания кустарничков и сосны. Таким образом, последовательность смен в ходе этой сукцессии всецело определяется постепенным накоплением торфа, образующегося из остатков отмерших растений. 111 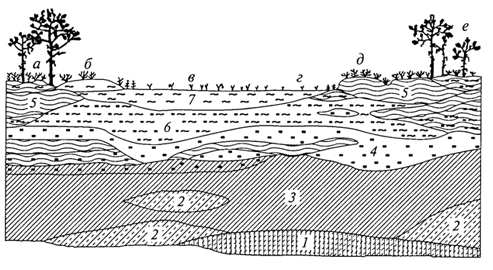 Рис. 17. Профиль среднетаежной болотной грядово-среднемочажинной экосистемы в Западной Сибири (О.Л.Лисе и др., 2002): Растительный покров: а -сосново-кустарничково-сфагново-зеленомошный; б- кустарничково-сфагновый; в -шейхцерево-пушицево-сфагновый; г -подбелово-сфагновый; д -клюквенно-сфагновый; е -сосново-кустарничково-сфагновый. Торфяная залежь: 1- ил; 2- камышовый торф; 3- осоковый торф; 4 - кустарничково-сфагновая; 5 - лесной торф; 6 -более старый сфагновый торф; 7- более молодой сфагновый торф. В центре болота заполненное водой "окно" ("болотный глазок"); болото окаймляет заболоченный лес Общее направление смен в ходе описанной сукцессии, весьма типичной для лесной зоны, связано с изменением характера увлажнения - переходом от натечного или почвенно-грунтового увлажнения в начале сукцессии (выходы грунтовых вод на поверхность или повышение их уровня) к увлажнению поверхности болота за счет влаги атмосферных осадков - верховое болото. Общее обеспечение водой растений при этом не изменяется, но меняется режим - от проточного к застойному - и обеспеченность растений элементами минерального питания. На так называемой стадии атмосферного питания болота минеральные элементы доступны растениям только из влаги атмосферных осадков или оседающей с ними на поверхность болот пыли. Это олиготрофная стадия развития болота (олигос - бедный, трофность - питание), которая как раз и характеризуется доминированием различных видов сфагновых мхов - мощных торфообразователей. Разнообразие аллогенных смен чрезвычайно велико. Они идут под воздействием климатических изменений, в связи с развитием рельефообразующих процессов, засолением-рассолением почв, изменением почвенно-грунтовых условий на террасах рек при прекращении воздействия половодья, жизнедеятельностью различных групп животных и особенно в результате деятельности 112 человека. Антропогенные сукцессии по масштабу воздействия и площади распространения стоят на первом месте в подавляющей части регионов суши. Живой покров некоторых природных зон, в частности степей и саванн, в значительной степени сформирован под воздействием выпаса и, по существу, представлен различными стадиями пастбищных (или пасквальных) сукцессии. Исследованиями Г. Н. Высоцкого и И. К. Пачоского еще в начале века были выявлены важнейшие закономерности развития пастбищной сукцессии в степях европейской части России. Так, И. К. Пачоский (1914) для типчаково-ковыльной подзоны выделил следующие стадии развития сукцессии. I. Стадия недостаточного выпаса (или полное отсутствие последнего). Характерно скопление большого количества отмерших надземных органов растений, что препятствует нормальному росту типичных степных трав. Семенное размножение дерновинных злаков подавлено. Места, освобождающиеся после отмирания старых дерновин, занимают главным образом корневищные злаки (пырей ползучий, костер безостый) и некоторые двулетние двудольные. Поскольку испарение с поверхности почвы сокращается, происходит мезофитизация травостоя. II. Стадия умеренного выпаса (ковыльная стадия, по Е.М.Лавренко). Все дерновинные злаки хорошо развиты, много молодых дерновин (показатель успешного семенного возобновления дерновинных злаков, одно- и двулетних растений). В этой стадии степь дает максимальную продукцию. III. Стадия угасания ковылей (типчаковая стадия, по Е. М.Лавренко). По мере увеличения интенсивности выпаса ковыли постепенно отмирают: вначале ковыль опушеннолистный, узколистный, украинский, затем ковыль Лессинга и, наконец, ковыль-волосатик - тырса; начинает преобладать типчак, в дальнейшем образующий сплошной покров. Продуктивность снижается, растительность становится более ксерофитной. Типчаковые степи, представляющие определенную стадию пастбищной дигрессии типчаково-ковыльных степей, широко распространены. IV. Стадия тонконогого сбоя. При усилении выпаса типчак изреживается, начинает преобладать хорошо переносящий вытаптывание мятлик луковичный. V. Стадия сбоя (по И. К. Пачоскому, стадия выгона) - наиболее сильно вытаптываемые места (стойбища, близ селений, колодцев), преобладают однолетники со стелющимися побегами (лебеда татарская, рогач песчаный, спорыш и др.). По мере перестройки и ксерофитизации растительности происходит расселение и увеличение численности более сухолюбивых животных - насекомых, птиц, млекопитающих (суслики, тушканчики, степные виды полевок). Возрастает численность специфических 113 паразитов домашнего скота, некоторых крупных хищников, происходит концентрация падалыциков. Выпас домашнего скота в лесах также приводит к сменам; под влиянием вытаптывания и стравливания меняется не только травяной покров, но часто прекращается и естественное возобновление древесных пород, что происходит как за счет непосредственного уничтожения молодых деревьев скотом, так и в силу уплотнения верхних слоев почвы. Уплотнение этого слоя почвы, перенасыщенной влагой, часто приводит к формированию своеобразного микрорельефа - скотобойных кочек (зоогенная мозаичность). В сухих и умеренно влажных условиях часто наблюдается иссушение почв. Интенсивное вытаптывание, особенно в засушливых климатах, на легких по механическому составу почвах вызывает развитие эоловых процессов, что может привести к формированию бугристых песков, почти лишенных растительности. Задолго до того, как появились стада домашнего скота и человек перешел к скотоводству, растительный покров находился под воздействием животных. Наиболее хорошо изучена роль животных в формировании степных и пустынных экосистем. Роющая деятельность сурков, сусликов, песчанок и других грызунов вызывает смены растительности на нарушенных участках, часто определяя ее состояние (А.Н.Формозов, А.Г.Воронов, 1939). Воздействие копытных, сформировавшееся в процессе естественной эволюции, специфично и направлено на поддержание устойчивости растительных сообществ. Известно, что в африканских саваннах большие стада многих видов копытных не разрушают растительность благодаря выработанным в ходе эволюции сложным взаимоотношениям. Разные виды животных скусывают растения на неодинаковой высоте, предпочитают разные корма и, передвигаясь в течение суток, охватывают большие площади, не сосредоточиваясь на отдельных участках, как домашний скот. Не менее значимы сукцессии, возникающие под воздействием пожаров, пирогенные (пирос - огонь) смены, довольно хорошо изученные в лесах. Так, низовой пожар, прошедший в сосновом бору-зеленомошнике или беломошнике, уничтожает главным образом напочвенный покров. Под сохранившимся пологом сосны разрастаются травы, преимущественно злаки. В случае сильного верхового пожара, который уничтожает полог сосны или ели, возникают совершенно иные условия и развивается длительная, складывающаяся из нескольких стадий сукцессия. На открытых участках, где почва не защищена от инсоляции, хорошо разрастаются светолюбивые травянистые растения, малина, а потом и такие древесные породы, как береза или осина, обыкновенно обильно плодоносящие, с легкими семенами, распространяемыми ветром. По мере смыкания крон березы или осины образуется 114 сплошной полог, под которым постепенно снова поселяется ель, формируя новое сообщество. В то же время с ростом и развитием хвойных пород, особенно после того как они выйдут в первый ярус древостоя, возобновление осины или березы под их пологом прекращается. Часто меньше, чем за 100 лет, прослеживаются стадии восстановления прежних сосновых или еловых лесов. Аналогично протекают и сукцессии на вырубках. Параллельно происходят смены животного населения: на ранних, так называемых открытых стадиях сукцессии характерны разнообразные насекомые, грызуны, свойственные лугам и лесным полянам, большое число видов птиц. С развитием кустарников, преимущественно ягодных (малины, жимолости и др.), вырубки охотно посещают медведи; ягоды потребляют и рыжие полевки, численность которых резко увеличивается; находит обильный корм на вырубках тетерев. На стадии развития мелколиственных пород (березы и осины) на вырубках охотно кормятся лоси и зайцы, сюда вселяются специфические для мелколиственных пород растительноядные насекомые. Наконец, под пологом мелколиственного леса развивается молодняк коренных хвойных пород и состав обитателей леса приближается к исходному. Рассматриваемые сукцессии столь распространены, что почти все березняки и осинники в таежной зоне представляют собой стадии, развившиеся на месте еловых, пихтовых, лиственничных лесов. С момента овладения огнем человек широко использовал пожары (так называемые палы) для расширения и поддержания продуктивности биоценозов степей, прерий, саванн. В различных регионах тропического пояса пожары и в настоящее время - важнейший способ регулирования состояния пастбищ, метод борьбы с разрастанием кустарников, удаления сухих частей растений в сухой сезон. Воздействие пожаров в эволюционном масштабе времени привело к тому, что в разных типах биоценозов в различных регионах суши формировались виды, в той или иной степени устойчивые к их воздействию, что определило их место в пирогенных сукцессиях. Например, многие виды семейства протейных в Австралии распространяются только после воздействия огня на их плоды. Восстановление биоценозов после нарушения демонстрируют сукцессии на залежах. При зарастании распаханной целинной степи вначале появляются однолетние, двух- и трехлетние травянистые растения, обладающие летучими семенами и отличающиеся быстрым ростом. Они образуют неустойчивое сообщество, сменяемое корневищными злаками (пырей и др.), образующими сплошной покров. В это сообщество внедряются дерновинные злаки (ковыль, типчак, тонконог); их разрастание приводит к восстановлению типичного степного фитоценоза. 115 В связи с эколого-биологическими особенностями и разными стратегиями жизни возникло представление о ранне- и позднесукцессионных видах. Как правило, жизненный цикл раннесукцессионных видов короток, они характеризуются быстрым ростом и высокой способностью к поселению на нарушенных участках. Для позднесукцессионных видов свойственны более низкая скорость роста, большая продолжительность жизни, значительно меньшая семенная продукция. В табл. 2 приведены важнейшие характеристики растений, свойственные ранним и поздним стадиям сукцессии. В ходе сукцессии изменяются продукция, биомасса, возрастает участие надземной биомассы растений в общей биомассе, в стадии развития кустарников или молодых лесов надземная биомасса в 2-3 раза превосходит массу корней, в спелых лесах это соотношение повышается до 5 - 6. Масса отмерших частей растений, так называемой подстилки, возрастает от 0,1 - 0,4 до 1,6 кг/м2 в молодых лесах, достигая еще большей величины в спелых древостоях. Не менее важная закономерность - изменение видового разнообразия на разных стадиях сукцессии (рис. 18). На более поздних Таблица 2 Характеристики растений, свойственные ранним и поздним стадиям сукцессии (по Р.Риклефс, 1979) 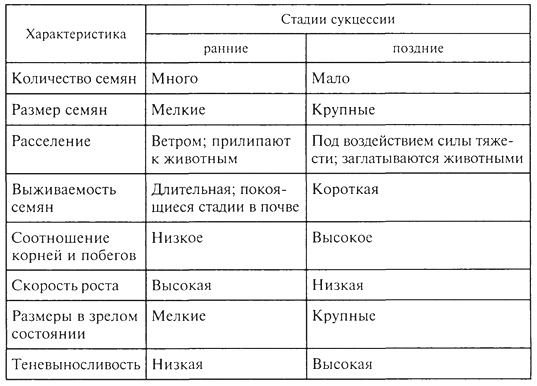 116 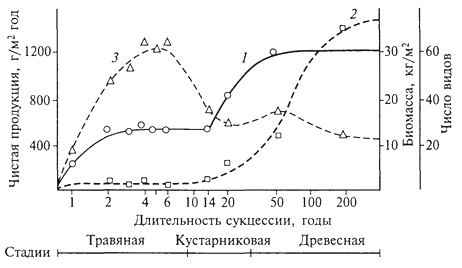 Рис. 18. Изменение чистой продукции (1), биомассы (2) и видового разнообразия (3) дубово-соснового леса (Брукхейвен, штат Нью-Йорк) в ходе сукцессии (Р.Уиттекер, 1980): 1 - чистая продукция увеличивается до стабильного уровня в травяной стадии, затем, поскольку в сообщества внедряются деревья, вновь возрастает до уровня, который может сохраняться в климаксе; 2- биомасса низка в травяных стадиях, но быстро увеличивается при накоплении древесных тканей кустарников и деревьев; климаксовый уровень биомассы, вероятно, может быть достигнут не ранее чем через 400 лет; 3 -видовое разнообразие (число видов на площади 0,3 га) возрастает в последних травяных стадиях, далее снижается в кустарниковых стадиях (14-20 лет), несколько увеличивается в молодом лесу (50 лет) и вновь уменьшается в климаксе (время дано в логарифмическом масштабе, что позволяет расширить ранние и сжать поздние стадии сукцессии) стадиях, связанных со становлением лесных биоценозов, по мере формирования структуры и смыкания лесного полога число видов часто убывает, как это происходит на стадии формирования ельников, дубрав, где под пологом уживается сравнительно ограниченное число видов. В ходе сукцессии происходит усложнение структуры биоценоза, ярусное расчленение. Скорость (время) развития от стадии к стадии в ходе сукцессии замедляется, что связано со сменой небольших и короткоживущих растений первых стадий более крупными и длительно живущими - на поздних. Так, широко известная и описанная выше смена после пожара в таежной зоне демонстрирует переход от одно- и многолетних травянистых растений к относительно недолговечным березе или осине, под пологом которых начинает развиваться ель, порода, длительно живущая. Биоценозы, сформированные долгоживущими породами, характеризуются относительной устойчивостью, как правило, на этих стадиях уже не происходит внедрения новых видов. Первым стадиям присуще, по определению Л. Г. Раменского, раздельно-зарослевое сложение, когда виды, эксплеренты по своей 117 жизненной стратегии, образуют несомкнутые, иногда почти одновидовые заросли; здесь много видов с легко распространяющимися семенами, сорняков, вегетативно подвижных растений (с ползучими корневищами, отпрысками, плетями). Для следующих стадий характерно смешанно-пятнистое сложение, когда выпадают почти все малолетние и многие случайные растения, - это простые группировки. Далее на более поздних стадиях происходит формирование сложения, свойственного данному биоценозу, с образованием ярусной структуры, свойственных ему микроценозов и устойчивого видового состава. 118 УСТОЙЧИВОСТЬ БИОЦЕНОЗОВ Состав и структура биоценоза в ходе сукцессии претерпевают глубокие изменения, которые, однако, не беспредельны. Как было показано выше, и при восстановлении близкого к исходному биоценоза (вторичные сукцессии), и при освоении биотой незаселенных ранее субстратов (первичные сукцессии) на последних стадиях происходит замедление смен, биоценозы приобретают относительную устойчивость. По отношению к таким сообществам употребляется термин "климакс", введенный в литературу американским геоботаником и экологом Ф. Клементсом. Климакс-сообщества - относительно устойчивые, способные к самоподдержанию и самовосстановлению, формирующиеся на хорошо развитых почвах, характеризующиеся устойчивостью видового состава, структуры, известной амплитудой сезонной и разногодичной изменчивости. Устойчивость климаксов сочетается с локальными нарушениями (ветровалами, деятельностью землероев и т.п.), которые способствуют "обновлению" определенных структурных частей биоценоза. Так, спелые деревья легко подвергаются ветровалу, что сопровождается осветлением, нарушением почвы; в "окнах" создаются условия, благоприятные для появления всходов, а затем и нового поколения деревьев. Часто при этом наблюдается смена пород: на осветленных участках в таежных лесах могут развиться береза, осина, сосна или рябина, под пологом которых начнется возобновление основных пород. Имеются данные (Р. Уиттекер, 1980) по развитию сукцессии на песчаных дюнах побережья оз. Мичиган, где для достижения климаксовой стадии дубового леса потребовалась почти тысяча лет. На залежах в степной зоне для достижения этой стадии требуется порядка 100 - 300 лет. Высока вероятность возникновения в течение такого времени ураганов, пожаров и других катастрофических явлений, которые могут "вернуть" сукцессию на начальные этапы ее развития. Несомненно, в климатических условиях, близких к экстремальным, время достижения климаксовой стадии 118 возрастает. Так, для тундры приводится время порядка 5000 лет. В течение таких больших временных отрезков происходят существенные климатические изменения, что может замедлить или приостановить развитие климаксовой стадии. Вероятность достижения климакса во многом зависит от деятельности человека, часто прерывающего поступательный ход сукцессии. В современной структуре живого покрова различных регионов суши практически не сохранилось сколько-нибудь обширных площадей, которые не испытали бы антропогенного воздействия той или иной степени интенсивности и повторяемости. В то же время для решения широкого крута задач, связанных с разработкой стратегии природопользования и сохранения биоразнообразия, необходимо представление о наименее нарушенных и наиболее стабильных биоценозах, свойственных тому или иному региону. В связи с этим особенно большое значение приобретают исследования в тех районах, где отсутствовали крупные нарушения и живой покров в наибольшей степени сохранил свое первоначальное состояние, которое находится в динамическом равновесии с современными природными условиями. Так, для Средней Европы, где практически не осталось ненарушенных лесов, известное представление о структуре и составе близких к климаксу биоценозов дает один из лесных массивов в Австрии, который никогда не подвергался рубке и использовался только как охотничье угодье. Характерна смешанная структура древостоя - бук, пихта, ель, причем бук входит во второй древесный ярус. Древостой неоднороден, бук и пихта часто произрастают куртинами. Высота древостоя 35 - 40 м, отдельных, наиболее старых пихт - 50 м при диаметре ствола в один метр. Хорошо развит подрост бука и ели, подрост пихты сильно поедается копытными. В травяном покрове характерно сочетание видов, с одной стороны, присущих широколиственным лесам (ясменник душистый, зеленчук, пролесник многолетний и др.), с другой - видов, преимущественно распространенных в хвойных лесах (черника, плаун годичный, марьянник лесной). В сложении травяного покрова выражена мелкая мозаичность, чередование пятен черники, плауна, пролесника, которые весьма нестабильны во времени. Черника и плаун появляются только на прогалах, где поздно стаивает скапливающийся за зиму снег; как только "окна" закрываются вновь сомкнувшимися кронами, черника и плаун здесь исчезают, но разрастаются на вновь появившихся прогалах. Некоторые виды, хорошо реагирующие на кислые почвы, формируют относительно временные поселения на рухляке отмершей древесины. В лесу можно обнаружить участки как бы выборочной рубки. Там, где произошло выпадение из состава древостоя наиболее старых деревьев, характерен полог более молодых древесных растений. Таким образом, в структуре девственного леса, относительно близкого к климаксу 119 в пространстве и во времени, происходит чередование древостоев и пятен травянистых растений. Исследования на таких эталонных участках позволяют не только изучить важнейшие закономерности самоподдержания климаксовых биоценозов, но и разработать принципы так называемого естественного лесоводства, когда взамен одновозрастных чистых насаждений воссоздаются разновозрастные и смешанные. Такие насаждения, как это показано на примере Германии, более устойчивы к колебаниям природных режимов, менее повреждаются насекомыми (которые предпочитают одну, вполне определенную породу) и грибковыми заболеваниями (Г.Вальтер, 1980). 120 |