Биогеография. Биогеография Г. М. Абдурахманов, Д. А. Криволуцкий, Е. Г. Мяло, Г. Н. Огуреева. Серия Высшее образование. М. Академия, 2003. 480 с. Isbn 5769509767 От издателя
 Скачать 13.83 Mb. Скачать 13.83 Mb.
|

|
ВЕРТИКАЛЬНАЯ СТРУКТУРА Вертикальная структура биоценоза определяется главным образом растениями, с высотой которых и их распределением по 96 группам высот связана дифференциация условий внутри биоценоза, от чего зависит распределение животных. Вертикальная структура биоценоза - одна из наиболее ярких черт его пространственной структуры. Важнейший фактор высотной дифференциации - интенсивность света, состав спектра. В любом биоценозе интенсивность света уменьшается сверху вниз за счет его поглощения: виды, относящиеся к разным жизненным формам и адаптированные к той или иной интенсивности света, размещаются на разной высоте. Расчленение биоценозов в их надземной части по вертикали на хорошо отграниченные горизонты - ярусы - наиболее четко проявляется в лесах умеренного пояса. Развитие представлений о ярусности лесных фитоценозов связано с работами исследователей скандинавской школы в лесах севера Финляндии. Значительный вклад в понимание ярусности внесли Г.Ф.Морозов и В.Н.Сукачев. В дальнейшем сформировалось представление о ярусах как о структурных частях биоценоза, каждый из которых по-разному использует и изменяет среду. В лесах умеренного пояса, образованных растениями, относящимися к разным жизненным формам, - деревьями, кустарниками, травами и кустарничками, мхами и лишайниками - обычно различают четыре яруса. Это древесный, кустарниковый, травяной или травяно-кустарничковый, моховой или мохово-лишайниковый ярусы (последние иногда называют напочвенным покровом). В зависимости от числа видов деревьев и их различий в высоте можно различать два и более древесных яруса. Кустарниковый ярус часто называют подлеском и включают в него кустарники и деревья, приобретающие в данном биоценозе кустарниковую форму. Так, В. Н. Сукачев включает в подлесок липу в подзоне южной тайги, где она часто принимает кустарниковую форму. В биоценозах тундры, где в жестких экстремальных условиях происходит "миниатюризация" жизни и растительный покров сжат по вертикали до дециметров и даже сантиметров, в одном ярусе оказываются растения разных жизненных форм; здесь выделяются мохово-кустарничковые, травяно-моховые, лишайниково-кустарничковые ярусы. Иными словами, жесткость условий среды определяет размещение на одном высотном уровне растений различных жизненных форм. Ярусность выражена и в травяных сообществах (лугах, степях, саваннах), но часто недостаточно отчетливо (рис. 12); в луговых и некоторых степных биоценозах расчленение на ярусы практически отсутствует. Помимо протяженности яруса по вертикали огромное значение имеет степень его сомкнутости. Сомкнутость древостоя обычно оценивается в долях единицы. Менее сомкнутые верхние ярусы определяют возможность развития более сомкнутых нижних. Это говорит о взаимовлиянии ярусов, об их взаимодействии 97  Рис. 12. Вертикальная проекция травостоя остепненного луга в Орловской области (Т. А. Работнов, 1968) внутри биоценоза и подтверждает положение о том, что ярус - это структурная часть биоценоза. Особое внимание исследователей привлекает структура влажного тропического леса. Это объясняется необычайным богатством форм в этом типе сообществ, его древностью и, следовательно, "слаженностью" жизни. Глубоко рассмотрел эти вопросы А. Г. Воронов (1985). Характерная особенность структуры влажного тропического леса - обилие числа видов деревьев. Как правило, наиболее многочисленные деревья относятся к невысоким, а самые высокие деревья представлены единичными экземплярами. Ярусы деревьев практически не выражены или выражены слабо, хотя некоторые авторы насчитывают в смешанном дождевом тропическом лесу пять ярусов, из них три древесных. Причинами невыраженности или слабой выраженности ярусов влажного тропического леса являются, во-первых, древность этого сообщества, в котором с максимальной полнотой используется пространство, а во-вторых, оптимальность условий, делающих возможным столь мощное развитие древостоя. Обилие разновозрастных стволов различной высоты окончательно затушевывает распределение растений 98 по ярусам. Следует полагать, что вообще дифференциация деревьев в сообществах по ярусам - явление вторичное, результат либо аридизации климата, либо понижения температуры (рис. 13). В тех случаях, когда ярусы образованы долгоживущими видами растении, они устойчивы по сезонам и годам. Например, ярус ели устойчив от сезона к сезону и не меняется по годам. Ярусы, образованные листопадными древесными породами, кустарниками и кустарничками, устойчивы по годам, но степень их сомкнутости варьируется от сезона к сезону, что определяет изменение условий для нижних ярусов в разные сезоны. Травяные ярусы, образованные вегетирующими большую часть или весь вегетационный период растениями, отмирающими в надземной части в конце его, непостоянны по сезонам и изменяются по годам. Наконец, эфемерные ярусы, образованные геофитами и терофитами, как правило, существуют часть сезона и обнаруживают большое варьирование по годам. Известное место в биоценозах занимают так называемые межъярусные растения - лианы и эпифиты Эпифиты поселяются на разной высоте на различных растениях, чаще всего на деревьях, используя их как опору и развивая корни в воздушной среде. Лианы, обвиваясь вокруг стволов деревьев, поднимаются в наиболее благоприятные для них условия освещенности. Доля этих растений 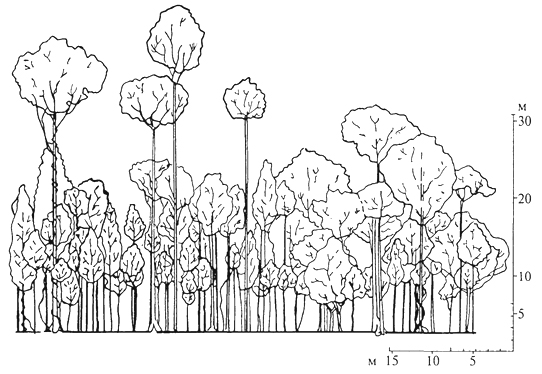 Рис. 13. Профильная диаграмма тропического леса на острове Калимантан, полоса леса длиной около 60 и шириной 8 м, деревья высотой более 7 м (П. Ричарде, 1960) 99 велика в различных типах тропических лесов, однако представлены они и в биоценозах умеренного пояса. Всем известны лишайники-эпифиты на стволах, столь свойственные таежным биоценозам, хмель, красиво цветущие лианы из рода клематис и др. Каждый вид в биоценозе представлен разновозрастной популяцией. Сложен вопрос о ярусной принадлежности всходов и молодых растений, не достигших нормальной для данных условий высоты и не вступивших еще в генеративный период, тех растений, которые часто объединяют в категорию подрост. Можно относить  Рис. 14. Подземная ярусность степного сообщества (А. М.Семенова-Тян-Шанская, 1950): 1 - житняк; 2 -ковыль; 3 -типчак; 4- полынь; 5 - кермек 100 подрост к ярусу, куда входят зрелые особи, трактуя в этом случае подрост как временный полог - в перспективе молодые растения войдут в этот ярус. Согласно другому подходу, подрост следует включать в тот ярус, где размещены его органы ассимиляции; в этом случае подрост часто оказывается в одном ярусе с подлеском. При этом один и тот же вид оказывается в разных ярусах, в зависимости от того, в какой ярус попадают разновозрастные особи вида. Таким образом, при рассмотрении вертикальной структуры биоценоза следует выделять степень расчленения биоценоза на ярусы, состав ярусов и их сформированность. Помимо надземной различают и подземную ярусность. Распределение корневых систем в подземной сфере обусловлено изменением с глубиной обеспеченности растений доступными формами элементов минерального питания, степени увлажнения почвы, а также ее аэрации. Установлено, что за очень редким исключением общая масса подземных органов растений закономерно снижается сверху вниз, особенно за счет мелких, в том числе сосущих, корней, в основном сосредоточенных в верхнем горизонте почвы. Даже глубоко укореняющиеся растения активно используют поверхностные горизонты почвы, в которых они формируют постоянно или временно существующие (эфемерные) корни. Постепенно уменьшаясь в количестве, корни многих растений проникают на значительную глубину, нередко существенно превышающую высоту надземных частей растений (рис. 14). Как было сказано выше, вертикальный каркас биоценоза определяется растениями. Наземные животные меняют свое ярусное положение в течение дня, года, жизни, проводя в том или ином ярусе более продолжительное время, чем в других. В большей степени связаны с определенными глубинами почвы различные населяющие ее беспозвоночные обитатели. Но и тут точнее говорить о тех градациях глубин, на которых встречаются представители той или иной систематической группы, того или иного вида, а не о приуроченности к определенному подземному ярусу. 101 ГОРИЗОНТАЛЬНАЯ СТРУКТУРА В горизонтальном сложении биоценозов проявляется мозаичность; практически в любом биоценозе могут быть выделены микроценозы (микрогруппировки, по терминологии некоторых авторов), различающиеся по видовому составу, количественному соотношению видов, сомкнутости и т.д. Самые простые примеры - осветленные межкроновые "пятна" под сомкнутым пологом леса; мощные (диаметром до нескольких десятков сантиметров) дерновины злаков (ковыля, например) в сухих степях и пятна почти 101 обнаженной поверхности почвы между дернинами. Мозаичность биоценозов формируется под воздействием как абиотических, так и биотических факторов. Степень однородности условий биотопа, в известной степени, относительна; так, нанорельеф - мельчайшие повышения и понижения - изменяет условия увлажнения и влияет на развитие всходов тех или иных растений. Сложная мозаика почвенных условий в пределах одного биотопа ярко обнаруживается там, где биоценозы формируются на твердых почвообразующих породах, различия в степени выветривания которых определяют размещение тех или иных видов. Огромную роль в формировании мозаичности играет жизнедеятельность самих растений и животных. Фитогенная мозаичность обязана своим возникновением средообразующей роли растений, само чередование подкроновых и межкроновых пятен в лесу - результат средообразующей (перехват света) жизнедеятельности растений. В смешанных хвойно-широколиственных лесах изучены различия в условиях, формирующихся, например, под дубом и под елью. Опад под елью состоит преимущественно из хвои, которая очень медленно разлагается, поэтому образуется мощная подстилка; в почвах, как правило, хорошо выражен подзолистый горизонт и слабо развит гумусовый. Опад дуба разлагается быстрее, в почвах гумусовый горизонт выражен лучше, а подзолистый хуже, чем под елью, прослеживается четкая приуроченность зеленых мхов, таежных трав и кустарничков (кислицы, черники и др.) к подкроновым пятнам ели и неморальных трав - к дубу (рис. 15). Степень выраженности мозаичности в различных лесных биоценозах варьирует в зависимости от мощности эдификаторов, степени сомкнутости верхнего яруса. Так, под густым пологом ели мозаичность может быть выражена очень слабо, в то время как в лиственничных лесах на северо-востоке России к подкроновым пятнам приурочены зеленые мхи, а в межкроновых осветленных участках развивается сплошной покров лишайников. Фитогенная мозаичность в травяных биоценозах выражена слабее, чем в лесных. Однако свойственное многим травянистым растениям групповое распределение (яркий пример - дерновины многих злаков, "пятна" и "латки" многих двудольных) также способствует формированию мозаичности. Часто мозаичность возникает за счет деятельности растений и абиотических факторов. Так, в пустынных биоценозах Монголии вокруг кустарников накапливается мелкозем, переносимый ветром, что ведет к образованию вокруг кустов караганы микроповышений, к которым приурочен определенный набор видов. Это так называемая эолово-фитогенная мозаичность. Зоогенная мозаичность, обусловленная жизнедеятельностью животных, чаще всего возникает в результате выброса на поверхность 102 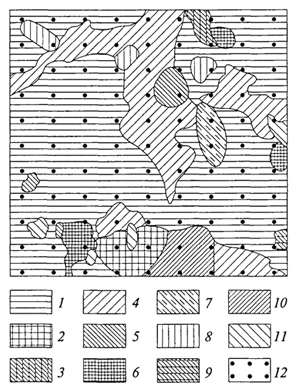 Рис. 15. Мозаичность в елово-дубовом лесу; микрогруппировки (Н.В.Дылис, 1969): 1 - елово-волосистоосоковая; 2- елово-кисличная; 3 -крупнопапоротниковая в "окне" древесного полога; 4- дубово-снытиевая; 5 - елово-костянично-пальчатоосоковая; 6 -елово-щитовниковая; 7 - дубово-осиново-зеленчуково-медуничная; 8 -березово-елово-мертвопокровная; 9 -лещиново-щитовниковая в "окне"; 10 -осиново-снытиевая; 11 - снытиевая в "окне"; 12 -маршрутные точки наблюдений кучек земли (кротовины в лесу и на лугах, сусликовины в степях), а также за счет интенсивного поедания растений вблизи нор, гнезд, различных убежищ. В возникновении мозаичности биоценозов умеренного пояса значительную роль играют муравьи, а биоценозов влажных тропиков - термиты. Микроценозы, микрогруппировки любого биоценоза находятся в тесной взаимозависимости друг от друга, связаны воедино переносом зачатков и продуктов жизнедеятельности, формируются в условиях свойственной данному биоценозу фитосреды, в первую очередь фитоклимата. 103 ИЗМЕНЧИВОСТЬ БИОЦЕНОЗОВ Динамичность - одно из фундаментальных свойств биоценозов и более сложных природных систем - биогеоценозов и экосистем. Как открытые системы, они характеризуются существенной 103 ролью вероятностных процессов в их функционировании и находятся в непрерывном изменении и развитии как под воздействием внешних условий, так и в связи с особенностями динамики численности и биологической активности отдельных популяций. Изменения биоценозов характеризуются суточной и сезонной ритмикой, могут проявляться в течение ряда лет, на протяжении столетий или же охватывать целые геологические эпохи, отражая развитие биосферы в целом. Суточные изменения биоценозов определяются суточной ритмикой жизнедеятельности организмов. Это колебания интенсивности транспирации, дыхания и фотосинтеза у растений; среди цветковых суточный ритм определяется и тем, что большинство видов открывает цветки днем. Характерна смена суточной активности у представителей различных групп животных (различают дневных, сумеречных и ночных). Сезонная изменчивость биоценозов проявляется в изменении состояния и активности видов, существенных колебаниях соотношения их численности в различные сезоны года. Важнейшие особенности сезонной изменчивости (ритмики) биоценозов связаны с сезонными изменениями светового и теплового режимов, а также условий увлажнения. Для большинства регионов Земли характерны изменения в течение года длительности фотопериода, интенсивности света, температуры, поступления атмосферных осадков. В процессе отбора видов на совместное обитание в данных климатических и почвенных условиях в состав биоценозов вошли виды, способные наиболее полно использовать условия, возникающие в отдельные сезоны года. Во влажном тропическом лесу, где в течение года не проявляется заметной дифференциации температурного режима и условий увлажнения, большая часть видов деревьев имеет существенно различные сезонные ритмы цветения и плодоношения; для одних характерно относительно короткое обильное цветение в определенное время года, другие - цветут менее обильно в течение длительного периода. С видами, характеризующимися разным ритмом цветения, связаны и различные опылители. Так, одни виды колибри ориентированы на кратковременно цветущие виды, охватывая обширные территории в их поисках, другие - посещают любые доступные цветки необильно цветущих видов. В более контрастных природных условиях, где в течение года меняются условия тепла или увлажнения, сезонная изменчивость биоценозов выражена более отчетливо; важное значение имеет дифференциация на два основных отрезка - период активного развития видов (лето, влажный период - в тропическом поясе) и время их полного или хотя бы частичного покоя: для многих растений полупокой, для животных спячка (зима, сухой период). В неблагоприятные сезоны года многие виды животных мигрируют, например перелетные птицы, многие виды млекопитающих. 104 В целом от полярных до тропических широт длительность активного сезона изменяется от нескольких недель до года. Самые яркие сезонные изменения связаны с жизнедеятельностью растений и их эколого-биологическими особенностями. В зависимости от выраженности ярусности и сезонной устойчивости надземных органов растений находится и сезонная изменчивость структуры биоценоза. Так, например, в сосняке-беломошнике (с плотным напочвенным покровом кустистых лишайников) в обоих ярусах в течение всего года не происходит существенных изменений. В дубраве же от осени к весне происходят изменения, связанные с листопадом; весной в нижнем ярусе ярко проявляется ценотически обусловленная сезонная изменчивость структуры - развивается группа раннецветущих видов. Чаще всего объясняют сезонное изменение структуры в дубраве изменением условий освещенности под пологом дубового леса ранней весной, когда листья дуба еще не распустились, а температура воздуха и почвы уже положительная. Представление о смене состояний видов и изменении их роли в течение года дают фенологические спектры, составляемые на основании детальных многоразовых наблюдений в одних и тех же типах фитоценозов. В связи с сезонными изменениями биоценозов возникло понятие "смена аспектов" - изменение внешнего облика биоценозов в течение года или благоприятного для жизни периода. Во многих типах биоценозов важную роль в формировании аспектов играют сменяющиеся фазы цветения или плодоношения некоторых видов растений. Так, для степных биоценозов характерен ярко выраженный аспект плодоносящих видов ковыля или ранней фазы цветения прострела. Смена аспектов в той или иной мере свойственна почти всем биоценозам, но степень ее выраженности зависит от эколого-биологических особенностей видов и структуры биоценоза. Во многих типах темнохвойных лесов она может в той или иной мере проявляться только в травяном ярусе. В листопадных лесах смена аспектов проявляется во всех ярусах, будучи наиболее ярко выраженной в травяном. Наиболее четко смена аспектов проявляется в травяных многовидовых биоценозах; классический пример - смена аспектов в луговых степях, изучение которых связано с именем В. В.Алехина. В тех случаях, когда изменения режима тепла и/или увлажнения особенно контрастны, многие, а иногда почти все виды, формировавшие один аспект, исчезают из состава травостоя, сохраняясь в почве в виде жизнеспособных зачатков и/или семян. Так, в биоценозах южных пустынь Средней Азии весной формируется сомкнутый покров из осоки толстостолбиковой и мятлика луковичного, видов, полностью исчезающих из наземного покрова 105 по мере наступления сухого жаркого периода. Не менее контрастна смена аспектов в тех случаях, когда на сезонные климатические изменения накладываются значительные колебания гидрологического режима. В столь характерных для южных степей и полупустынь многочисленных западинах, известных в разных регионах под названием поды, лиманы, весной в условиях поверхностного затопления развиваются частуха подорожниковая, ситняг болотный, иногда камыш озерный и некоторые другие водно-болотные и прибрежно-водные виды. После спада воды аспект создают пырей ползучий, некоторые виды разнотравья, а все гелофиты сохраняются только в почве в виде луковиц, клубней, корневищ. Таким образом, в разные отрезки вегетационного периода в один биоценоз входят экологически контрастные, в других условиях несовместимые виды. Сезонная изменчивость проявляется и в сменах животного населения. В летнее время для биоценозов тундры характерен большой набор видов птиц, насекомых и других животных. Зимой большинство птиц и северные олени мигрируют на юг, другие животные впадают в оцепенение и выключаются из активной жизни. Циклические (ритмические) изменения сообществ, происходящие от года к году или в течение нескольких лет, известны всем пользователям природных ресурсов и являются предметом специальных исследований. По отношению к этой изменчивости употребляются различные термины, от многолетней, погодичной, разногодичной изменчивости, которые предлагались разными исследователями, до получившего в последнее время наиболее широкое распространение термина "флуктуация". Многолетняя изменчивость, флуктуации, - одна из ярких способностей биоценоза реагировать на колебания параметров внешней среды изменением численности, соотношением популяций разных видов, изменением их жизненного состояния. Флуктуации биоценоза связаны также и с проявлением жизненных ритмов организмов, особенностями жизненного цикла различных видов, в первую очередь доминантов и эдификаторов, с особенностями динамики численности животных, массовым размножением патогенных организмов. Различия по годам в тепле и влаге, связанные с метеорологическими флуктуациями, прямо (для растений) или косвенно (для многих животных) оказывают влияние на численность популяций различных видов. В первую очередь происходят изменения продуктивности растений, прироста, урожайности, семенной продукции. В известной связи с этим находятся колебания численности многих видов-фитофагов (насекомых, мышевидных грызунов); существует определенная связь между колебаниями их численности и численностью хищников (рис. 16). 106 В степях Монголии в годы массового размножения полевки Брандта (одного из типичных мышевидных грызунов) происходит разрыхление почвы, увеличение содержания в почве азота за счет минерализации отмерших растений, экскрементов животных. Разрыхление почвы на значительных площадях приводит к разрастанию корневищных злаков, в первую очередь востреца, и почти полному отмиранию типичных для этих степей дерновинных злаков. Вострецовые травостои господствуют в течение нескольких лет, пока в результате снижения численности полевки не восстанавливается вновь господство дерновинных злаков. По многолетним наблюдениям в Барабинской степи выявлены интересные особенности флуктуации биоценозов в связи с чередованием нескольких засушливых и влажных лет подряд. Постепенное понижение уровня грунтовых вод в засушливый период 1950- 1955 гг. стимулировало проникновение вслед за ними корневых систем глубокоукореняющихся трав, например колосняка мохнатого, кермека Гмелина. У этих видов отсутствовала реакция на иссушение почвы, но затем, при повышении уровня грунтовых вод, они пострадали в первую очередь из-за отмирания затопленных водой корневых окончаний. Засуха особенно сильно сказалась на влаголюбивых видах, которые или гибли, изреживаясь, или переходили в состояние покоя. Усилившееся в засуху засоление почв послужило причиной массового отмирания некоторых типичных степных растений. Во влажные годы, когда промачивание почвы привело к ее опреснению в верхних слоях, эти виды появились вновь. Высокая температура и усиление аэрации почвы способствовали развитию бактерий-нитрификаторов и накоплению нитратов, что стимулировало массовое размножение осота полевого, одуванчика и других азотолюбивых видов. Когда энергия нитрификации вслед за увеличением влажности почв снизилась,  Рис. 16. Изменение численности зайца-беляка (1) и рыси (2) в Республике Коми (С.П.Наумов, 1947) 107 эти виды, не выдержав конкуренции со злаками, перешли в угнетенное состояние. Изменения в биоценозах были усилены при массовом размножении в 1953 г. нестадных саранчовых. Прямокрылые съели до 80 - 90% массы мягколистных злаков: пырея, костра, типчака и др. У жестколистных колосняка, тростника и других они уничтожали всходы по мере их появления. В этот год бурно разрослось непоедаемое саранчовыми разнотравье, для которого была устранена конкуренция со стороны злаков. Последствия размножения саранчовых сказались и на соотношении злаков в следующие два года. В засушливые годы муравьи заселили кроме возвышенностей высохшие низинные луга, что повлияло и на условия произрастания растений (К. А. Куркин, И. В.Стебаев, 1976). Такие флуктуации имеют тенденцию повторяться вслед за периодическими колебаниями климата, обусловленными изменением общей циркуляции атмосферы, усилением и ослаблением солнечной активности. На фоне колебаний природных режимов в ходе флуктуации сохраняется в основном видовой состав при существенном варьировании численности, изменении ценотической значимости различных видов. Малочисленные виды в определенных ситуациях увеличивают обилие при одновременном понижении численности других видов. В некоторых случаях при этом происходят смены доминантов. "Исчезновение" и появление каких-то видов в большинстве случаев - кажущееся. В неблагоприятные для некоторых растений годы они переходят в угнетенное или покоящееся состояние. Таким образом, распространяя данное Т. А. Работновым (1983) определение флуктуации фитоценозов на биоценозы, их можно определить как ненаправленные, различно ориентированные или циклические изменения биоценозов от года к году или в течение краткосрочных климатических циклов, завершающихся возвратом к исходному или близкому к нему состоянию. 108 |