Биогеография. Биогеография Г. М. Абдурахманов, Д. А. Криволуцкий, Е. Г. Мяло, Г. Н. Огуреева. Серия Высшее образование. М. Академия, 2003. 480 с. Isbn 5769509767 От издателя
 Скачать 13.83 Mb. Скачать 13.83 Mb.
|

|
ГЛАВА IV СТРУКТУРА ЖИВОГО ПОКРОВА СУШИ Непрерывность биоценотического покрова - континуум. Непрерывность живого покрова, или континуум, - фундаментальная черта организации биосферы. Так, растительность одевает сушу в виде сплошного покрова, при этом в почве корни растущих рядом особей переплетаются, а в наземных частях листва и ветви перекрывают друг друга, создавая "непрерывную ткань взаимовлияния". Впервые представление о непрерывности растительного покрова было сформулировано выдающимся русским ученым Л. Г. Раменским, аналогичные положения были обоснованы в США в работах Г. Глизона. Согласно этим представлениям, растительный покров - непрерывно меняющееся в пространстве сочетание популяций видов: разные виды, произрастающие совместно в различных комбинациях, образуют плавно переходящие друг в друга меняющиеся сочетания. На местности нет ясно отграниченных друг от друга фитоценозов (топографический континуум), как нет и четко очерченных фитоценозов в ходе развития сукцессии - переходы от одной комбинации совместно произрастающих видов к другой во времени плавны и непрерывны (временной континуум). В ходе сукцессии на гарях и вырубках в таежных регионах происходит смена березовых (одна из стадий сукцессии) лесов еловыми. Березняки и ельники - биоценозы, хорошо отличающиеся друг от друга видовым составом и структурой. Однако в течение длительного времени, пока ель, постепенно поселяясь и развиваясь под пологом березы, не выйдет в первый ярус и не произойдет отмирание березы, трудно отделить березняки от ельников. Также постепенно во времени происходит смена животного населения, что определяет непрерывность, плавность смены и биоценозов. Л. Г. Раменским (1924) впервые были разработаны методы изучения непрерывного распределения растений в связи с действием отдельных или группы экологических факторов, получившие впоследствии название градиентного анализа. Используя эти методы, исследования растительности или биоценозов проводят через определенные интервалы градиента условий существования, например по комплексному градиенту абсолютной высоты при подъеме в горы через 100 м или по градиенту глубины снежного покрова. Колебания численности (обилия) организмов вдоль ряда градиентов среды описываются колоколообразными кривыми, которые 121 имеют тенденцию к изменению независимо друг от друга и часто широко перекрываются. Это показатель того, что популяция каждого вида предъявляет специфические требования к среде, избирательна по отношению к условиям местообитания, имеет и собственные области доминирования. Непрерывное замещение одних видов другими вдоль градиента условий среды - континуум. Концепция континуума живого покрова базируется на двух положениях: представлении об экологической индивидуальности видов и непрерывности изменений факторов среды в пространстве и во времени. Растительный покров может быть представлен как структурный континуум, где выделяются участки, относительно однотипные по составу, структуре и иным признакам, и переходные, смешанные по составу комбинации видов, то более, то менее широкие. Резкость, выраженность переходной полосы зависит прежде всего от различий в условиях среды; хорошо выделяются "перерывы в непрерывности", например, при смене различных по составу и структуре почвообразующих пород. Прерывистость, резкий переход от одного однотипного участка к другому, может быть связана с существованием пороговых величин в непрерывно изменяющихся условиях среды с наличием переломных пунктов в изменении прямодействующих экологических режимов. Так, на рис. 19 представлено распределение растительности по градиенту глубины снежного покрова на Новосибирских островах. Как показала В.Д.Александрова (1969), здесь при глубине снега не более 12 см под действием прямого солнечного излучения подстилающая поверхность нагревается выше точки таяния льда еще при отрицательной температуре воздуха, вызывая образование снежных парничков за месяц и более до того, как начнет освобождаться от снега остальная поверхность тундры. При  Рис. 19. Пространственный континуум как следствие экологической индивидуальности видов; зависимость обилия видов в арктической тундре от глубины снежного покрова (В.Д.Александрова, 1965): 1 -Potentilla emarginata; 2 - Alopecurus alpinus; 3 - Papaver polare; 4-Draba pohle; 5 - Ranunculus nivalis; 6 - Cerastium bialynickii; 7 - Salix polaris; 8 - Deschampsia brevifolia; 9 - Androsace triflora; 10 - Juncus biglumis; 11 - Phippsia algida; 12 - Ranunculus pygmaeus 122 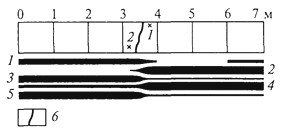 Рис. 20. Относительно резкая граница между фитоценозами соснового леса на песчаной дюне в районе Стокгольма (В.Д.Александрова, 1968): 1 - вереск (Calluna vulgaris); 2 -водяника (Empetrutn nigrum); 3 -лишайница оленья (Cladonia rangeferina); 4 -лишайница лесная (Cladonia sylvaticd); 5 -хилокомиум (Hylocomiumproliferum); 6- граница между фитоценозами; × - последние побеги вереска и водяники; ширина черных полос отражает степень обилия видов глубине снега более 12- 13 см такого эффекта уже не наблюдается - указанная мощность снегового покрова является границей смены доминирующих видов и самих фитоценозов. Как известно, экологические свойства видов в процессе эволюции формировались не в условиях индивидуального существования, а всегда в сообитании с другими видами. В результате сопряженной эволюции и дифференциации экологических ниш выработались сравнительно устойчивые сочетания видов, что также обусловливает выделение однотипных участков. Континуальность хорошо выражена в тех случаях, когда основу растительного покрова составляют вегетативно подвижные травянистые растения (яркий пример - луга). В лесах, при смене древесных пород-эдификаторов, как правило, возникают относительно четкие границы. В зависимости от характера смены условий (постепенной или резкой), мощности средообразующей деятельности организмов находится ширина переходной полосы между разными биоценозами и, следовательно, сам характер границы. Границы могут быть более или менее резкими и очень плавными (рис. 20). Особенно большое влияние на характер границ между биоценозами оказывает деятельность человека: границы лес - пашня, лес - луговая поляна в лесу всегда достаточно резки. Правда, и в этом случае есть переходная полоса - опушка леса на границе с пашней или лугом. В связи с этим возникло понятие экотон. Для экотона - переходного сообщества между двумя относительно четко различающимися сообществами - характерно участие видов, представителей обоих смежных биоценозов, но они также могут включать и виды, преимущественно свойственные экотонам, так называемые краевые виды. Часто экотоны характеризуются большим числом видов, чем смежные с ними. Известно, что на опушках леса растительность 123 пышнее и богаче, здесь гнездится больше видов птиц, встречается больше видов насекомых, пауков и т.д., чем в глубине леса. Это обусловлено более сильным варьированием условий освещенности, влажности, температуры на опушке, чем под пологом сомкнутого леса. Это так называемый пограничный, или "опушечный" эффект. В настоящее время понятие экотон стали применять более широко, часто распространяя его на целые зоны. Так, известный немецкий ботанико-географ и эколог Г. Вальтер (1976) рассматривает как экотоны лесостепь, лесотундру. Проявление пограничного эффекта прослеживается не только при анализе контактов двух фитоценозов (биоценозов), но и при анализе контактных полос между типами растительности или формациями. Исключительное видовое богатство флоры и фауны европейской лесостепи (как контактной полосы между широколиственными лесами и степями) объясняется, в том числе и ее экотонным характером. 124 КЛАССИФИКАЦИИ БИОЦЕНОЗОВ В природе существует бесконечное число биоценозов. Выявление закономерностей их географического размещения, важнейших особенностей структуры живого покрова того или иного региона требует их классификации, что необходимо и в практических целях. Классификация - это логическая операция, которая заключается в разбиении данного множества объектов на подмножества, или классы. Под классом понимается совокупность объектов, имеющих общие признаки, поэтому объекты данного класса отличаются от других объектов, которые этими признаками не обладают. В основе любой научной классификации лежит процедура выбора важнейших признаков, свойств, которые позволяют свести бесконечное множество биоценозов к какому-то ограниченному их числу. Сложность классификации биоценозов заключается в выборе признаков; можно классифицировать по видовому составу, по доминантам, по особенностям ярусной структуры с учетом доминантов, по набору жизненных форм или форм роста. Различные свойства, признаки сообществ, которые могут быть положены в основу классификации, не изменяются параллельно друг другу. Сообщества динамичны, что тоже необходимо учитывать при классификации. Не существует одного какого-либо "правильного" подхода к классификации, известно несколько таких систем. Чаще всего при классификации биоценозов используются классификации растительности как ведущего компонента их структуры и состава. В 1910 г. на 3-м Международном ботаническом конгрессе в Брюсселе принято решение считать основной типологической единицей 124 классификации растительности ассоциацию. Одно из лучших определений ассоциации принадлежит В.Н.Сукачеву (1957): "Ассоциация (тип фитоценоза) объединяет все фитоценозы, однородно участвующие в аккумуляции и трансформации вещества и энергии на поверхности земли, или, точнее, фитогеосферы. В соответствии с этим они характеризуются в основном однородным видовым составом, однородной структурой и однородным составом факторов среды, влияющих на фитоценологический процесс". Согласно одному из принятых подходов, в ассоциацию объединяются фитоценозы, сходные по видовому составу, при тождестве доминантов каждого яруса. Это определяет однородность структуры, высокое сходство экологических условий. Так, все фитоценозы, где доминантом первого яруса является ель, в кустарничковом ярусе доминирует черника, а в напочвенном покрове - зеленые мхи, объединяются в ассоциацию ельник-черничник- зеленомошник. Одинаковые доминанты и эдификаторы каждого яруса определяют экологию биоценоза, его влияние на все факторы внешней среды, с чем связано, в свою очередь, и сходство в подборе второстепенных видов. Выделение ассоциаций по доминантам сопряжено с трудностями там, где развита полидоминантность. Так, во влажных тропических лесах, где в древесном пологе представлено много видов с примерно равным участием, доминантный подход не применим. Полидоминантность свойственна многим травяным (луговым, степным) и некоторым другим биоценозам. Любая ассоциация характеризуется свойственной ей областью географического распространения. Это та территория, где сохраняется доминирование видов каждого яруса и, следовательно, структурные особенности биоценоза. Каждая ассоциация охватывает определенный спектр экологических условий, формирование которых связано с теми или иными почвенно-геоморфологическими условиями. Иными словами, ассоциации локализованы в ландшафтной структуре территории. Смена сообществ, относящихся к разным ассоциациям, лучше всего улавливается на экологических профилях, охватывающих разнообразие местообитаний по рельефу. В связи с тем что условия для доминирования видов определяются конкурентными взаимоотношениями, сообщества, относящиеся к одной ассоциации, в пределах области ее распространения распределены прерывисто. Цель классификации растительности - не только выявить таксоны одного ранга (ассоциацию), отражающие эколого-географические связи на топологическом или ландшафтном уровнях дифференциации биосферы. Таксономическая единица классификации растительности на региональном уровне - формация. К одной формации относятся ассоциации с одними и теми же доминантами и эдификаторами главного яруса. Таким образом, все ельники, 125 где доминантом главного яруса является ель европейская, объединяются в формацию ели европейской. Формация ели европейской объединяет ельники-зеленомошники (черничники, кисличники и др.), ельники-долгомошники (с напочвенным покровом из мхов рода политрихум), ельники сфагновые, ельники сложные, с подлеском из лещины, липы. Область распространения (ареал формации), учитывая относительно большое разнообразие экологических условий, где возможно доминирование данного вида в главном ярусе, охватывает обширные территории и широкий диапазон почвенных и климатических условий. Таксономическая единица самого высокого ранга - тип растительности. При ее выделении основное внимание обращается на принадлежность эдификаторов и доминантов к одной или сходным жизненным формам: например, хвойные леса умеренного пояса (светло- и темнохвойные); широколиственные леса с господством листопадных широколиственных пород (дуба, липы и др.); степи с господством многолетних микротермных ксерофильных травянистых растений (преимущественно дерновинных злаков). Поскольку в основу выделения типа растительности положено сходство в наборе жизненных форм, ареал типа растительности охватывает весь диапазон тех природных условий, в первую очередь климатических, в которых функциональны адаптации этой жизненной формы. Известное единство условий среды на территории распространения типа растительности проявляется в первую очередь в особенностях гидротермического режима, соотношения тепла и влаги, наличии или отсутствии перерыва в вегетации, обусловленного отрицательными температурами и/или засушливым сезоном. Ареалы типов растительности, как правило, достаточно велики, по их рубежам проводится зонально-подзональное деление растительного покрова. Существуют системы классификации, учитывающие преимущественно видовой состав (так называемые флористические) или строящиеся на особенностях местообитания. Последние известны как фитотопологические; при этом растительность подразделяют на основе различий в местоположении. Так, распространенное деление лугов на материковые (водораздельные) и поемные - пример фитотопологического подхода. В свою очередь, материковые луга дифференцируются на суходольные (формирующиеся на местообитаниях с атмосферным увлажнением) и низинные (где в режиме влагообеспеченности существенную роль играет и почвенно-грунтовое увлажнение). Известная классификация лесов украинско-белорусского Полесья П.С.Погребняка (1968) является фитоэкологической (см. рис. 10). Строится так называемая эдафическая сетка, состоящая из двух координат: одна из них характеризует увлажнение, другая - богатство почвы. По богатству почвы выделяются четыре группы типов местообитаний (трофотопов): 126 боры (наиболее бедные почвы); простые субори, сложные субори, дубравы (наиболее богатые почвы). В пределах каждого трофотопа на основе растительности отграничивают шесть типов, различающихся по увлажнению (гидротопов): очень сухие, сухие, свежие, влажные, сырые, болота. Таким образом, в этой классификации, по сути дела, представлены типы местообитаний, выделенные по свойственной им растительности. Прямые и косвенные связи животного мира суши с растениями очевидны, что легло в основу представлений о биоме. Американские экологи для разграничения и характеристики совокупности растительности и животного населения крупных регионов суши взяли за основу единицу растительности примерно ранга типа со связанным с ним комплексом позвоночных. Эти "растительно-животные формации" получили от Ф. Клементса и С. Шелфорда название биомов - единиц, характеризующихся жизненными формами и доминирующими видами, например биом прерий. В различных весьма удаленных друг от друга территориях могут формироваться физиономически, по набору жизненных форм сходные биомы, отличающиеся по систематическому составу входящих в них видов. Это типы биомов, например тип биомов степей и прерий, характеризующийся высоким сходством структуры и ритмики биоценозов и большими таксономическими различиями. Многочисленные работы по установлению соответствия животного населения растительным ассоциациям, определяющимся доминирующими видами, дают противоречивые результаты. Подвижность животных, смена местообитаний в зависимости от стадий развития, по сезонам года требуют дифференцированного подхода для разных групп животных. Очень подвижные животные - млекопитающие, птицы - больше связаны с формациями, чем с ассоциациями. Многие в соответствии с их образом жизни являются членами двух или нескольких сообществ, сменяя их в течение суток: нора находится в одном сообществе, а охотничья территория - в другом. В то же время эколого-географические связи отдельных групп животных оказываются шире или уже рамок классификационных единиц растительности. В связи с этим существуют отдельные подходы к классификации животного населения. 127 СТРУКТУРА ЖИВОГО ПОКРОВА Элементарной ячейкой дифференциации живого покрова суши является биогеоценоз в границах фитоценоза, который никогда не существует изолированно, а связан территориально и функционально со смежными. Результатом этого являются сочетания (комбинации) сообществ, образующие структуру живого покрова, 127 которую можно исследовать в разных масштабах: суши в целом, материков, крупных регионов и ландшафтов. Различия в структуре живого покрова суши обусловлены многообразием изменений в пространстве климатических, орографических, почвенно-литологических условий, определяющих значительное число вариантов сочетаний разных типов сообществ друг с другом в тех или иных регионах. Макроструктура растительного покрова суши определяется дифференциацией природной среды на планетарном уровне и находит отражение в его зональном и высотно-поясном расчленении. В основу представлений о зональной структуре живого покрова планеты легли установленные связи между растительностью крупных регионов суши и климатом. Для выявления этих связей используется разработанный немецкими экологами метод климадиаграмм, позволяющий выявлять характерные особенности того или иного климата, проводить его сравнение с климатами других географических регионов (Г.Вальтер, 1968). При составлении климадиаграмм за основу берут чаще всего соотношение между средней месячной температурой и количеством осадков как 1 : 2, иными словами, 10°С соответствует 20 мм осадков. При таком масштабе засушливая часть года характеризуется кривой осадков, расположенной ниже температурной кривой. Температурная кривая может служить показателем годового хода испарения, она отражает расход влаги, а кривая осадков - ее приход; обе кривые дают представление о характере водного баланса данной территории (рис. 21). 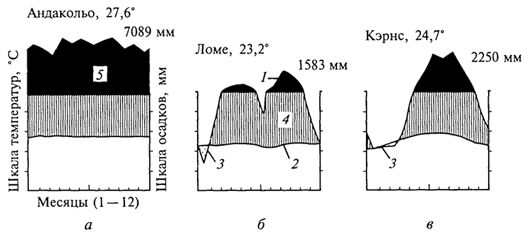 Рис. 21. Климадиаграммы тропического влажного дождевого леса (Г. Вальтер, 1968): а- в Колумбии; б- в Камеруне; в- в Австралии; 1 - кривые средних месячных осадков; 2 -кривые средних месячных значений температуры; 3 -слабо выраженный сухой период; 4-влажное время года; 5 - среднее месячное количество осадков, превышающее 100 мл 128 Соотношение температуры и осадков как 1 :2 применимо для территорий, где достаточно отчетливо выражен сухой период. Там, где период засухи просматривается слабо, используют соотношение 1:3, что позволяет выявить интенсивность и продолжительность полузасушливого периода. Используя климадиаграммы, можно выявить продолжительность вегетационного периода. Правда, термин "вегетационный период" требует уточнения. Это может быть и безморозный период, и время года со средней суточной температурой более 5, 10 или 15°С. В качестве показателя лучше всего использовать период со средней суточной температурой более 10°С, оцениваемой по кривым годового хода температуры воздуха. Важно также выявить на климадиаграммах продолжительность периода со средней суточной температурой ниже - 10°С, характеризующего термические условия очень холодных континентальных областей с суровыми зимами. Таким образом, климадиаграмма дает целостное представление о климате, что особенно важно при экологических исследованиях, когда требуется оценка совокупного воздействия различных элементов климата на биоту. Тесные связи растительности с климатом еще в начале XX в. впервые были использованы немецким биоклиматологом В.Кеппеном для классификации климатов по их отношению к растительности. Подразделяя растения на экологические группы по отношению к теплу и влаге, В. Кеппен выделял мегатермы (обитают при постоянно высоких температурах и влажности), ксерофилы (заселяют районы с сухим климатом, где хотя бы в отдельные сезоны отмечаются высокие температуры), мезотермы (обитатели умеренно теплых и умеренно влажных регионов с относительно теплой зимой), микротермы (произрастают в условиях низкой теплообеспеченности, обязательно с холодной зимой), гекистотермы (существуют в термических условиях холодных полярных стран). По климатическим показателям районов распространения этих экологических групп проведено биоклиматическое деление суши и выделены области распространения климатов мегатермических, ксерофильных, микротермических. Более дробную дифференциацию осуществляют по биоте. Так, среди мезотермических климатов выделяют климат оливки (средиземноморский, с сухим летом и влажной теплой зимой) и климат камелии (влажный субтропический с влажным летом и мягкой теплой зимой). Закономерности размещения классификационных единиц высокого ранга - типов растительности, типов биомов - в связи с изменением соотношения тепла и влаги по градиенту от низких широт к высоким выявляются при их размещении в осях влажности и температуры (рис. 22). В пределах тропических и субтропических областей, со средней температурой от 20 до 30 °С, типы 129 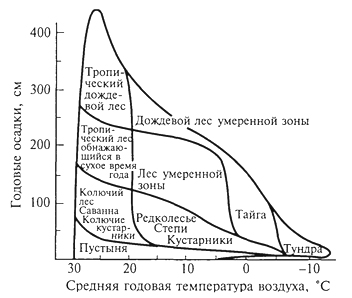 Рис. 22. Расположение основных типов биомов земного шара в осях влажности и температуры (Р. Риклефс, 1979) растительности изменяются от настоящего дождевого леса, где круглый год влажность высокая, до пустыни. Между этими "крайностями" располагаются влажные листопадные леса, где в сухое время года некоторые или все деревья теряют листву, а также сухие листопадные леса и/или кустарники, часто колючие. В умеренных широтах наблюдается смена растительности, биомов от лесов, развивающихся при достаточном увлажнении (дождевые леса умеренной зоны), к пустыням. Однако в арктических и субарктических широтах, где средняя годовая температура ниже -5 °С, размещается только один тип - тундра. Различия в условиях увлажнения в арктических и субарктических областях не оказывают существенного влияния. Влажная тундра перенасыщена водой, но, поскольку на протяжении большей части года эта вода остается замерзшей, а на глубине 50 - 60 см лежит слой вечной мерзлоты, она недоступна растениям (Р. Риклефс, 1979). На "сухом" конце градиента осадков в пределах каждой температурной зоны значительную роль в господстве тех или иных типов растительности играют пожары. В африканских саваннах и прериях на Среднем Западе США частые пожары уничтожают проростки деревьев и препятствуют возникновению лесов, условия для развития которых здесь вполне подходящие. Пожары благоприятствуют разрастанию многолетних травянистых растений, почки возобновления которых размещены ниже поверхности почвы и слабо или совсем не повреждаются огнем. Для выявления важнейших географических закономерностей размещения основных классификационных единиц живого покрова суши в биогеографии широко используются построения так называемого 130 идеального континента, или схемы, отражающей связь живого покрова с особенностями климата на идеально горизонтальной поверхности. Известная схема идеального континента К. Тролля (рис. 23) демонстрирует дифференциацию растительного покрова суши при отсутствии крупных горных поднятий и сохранении в принятом масштабе соотношения площади суша - океан на различных широтах. На схеме хорошо прослеживается зональность первого порядка фитосферы (согласно В. Б.Сочаве) - поясность живого покрова суши в связи с наиболее крупными различиями в биоклиматических условиях. Выделяют тропический, 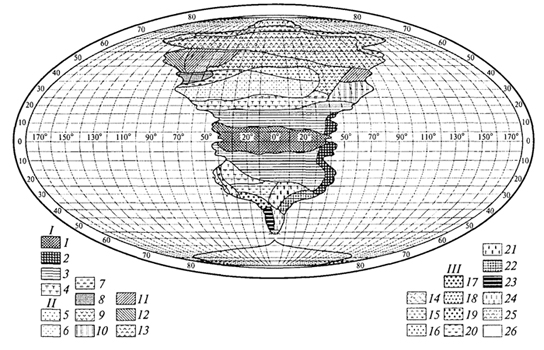 Рис. 23. Распределение растительности на "идеальном" континенте, составленное К.Троллем (Г.Вальтер, 1968): I. Тропические зоны: 1 -экваториальный дождевой лес; 2- тропические дождевые леса; 3 -тропические саванны (влажные саванны) и листопадные леса; 4 -тропические сухие саванны и колючие леса. II. Внетропические зоны северного полушария: 5-сухие пустыни; 6- холодные внутриматериковые пустыни; 7 - субтропические зимнезеленые степи; 8 -жестколистные леса и кустарники; 9 -зона степей с холодной зимой; 10- зона влажных муссонных и лавровых лесов; 11- летнезеленые леса; 12 - океанические летнезеленые листопадные и лавровые леса; 13 -бореальные хвойные леса; 14 -бореальные березовые леса; 15 - субарктические тундры; 16-высокоарктические тундры. III. Внетропические зоны южного полушария: 17 -прибрежные пустыни; 18 -пустыни, развитые в пределах пояса "Гаруа"; 19- жестколистные леса и кустарники; 20- субтропические колючие степи; 21- субтропические злаковники; 22- субтропические дождевые леса; 23 -умеренно холодные дождевые леса; 24 -степи Патагонии и Новой Зеландии; 25 -субантарктические туссоковые луга и болота; 26- антарктическая область материкового льда 131 внетропический северного полушария и южный внетропический пояса, рассматриваются зональные типы растительности, присущие каждому поясу. На схеме хорошо видны особенности макроструктуры растительного покрова, в частности приуроченность тех или иных зональных типов сообществ к определенным широтам, различие в наборе зон в восточных, западных и центральных частях континента. Обращает на себя внимание многообразие форм рисунков областей распространения зональных типов: наряду с зонами широтного простирания выделяются и зоны, вытянутые в субмеридиональном направлении, преимущественно в южном полушарии. Ряд четко выделяющихся в северном полушарии зон, протянувшихся через весь континент (бореальные хвойные леса, тундры и др.), в южном полушарии отсутствуют. На схеме "идеального" континента К. Тролля хорошо просматривается асимметрия типов растительности в южном и северном полушариях, видно, что макроструктура растительности южного полушария не является зеркальным отражением таковой северного полушария. Своеобразие растительного покрова южного полушария можно показать и на таком примере: южная граница распространения пальм (тропического семейства) пересекает Южный остров Новой Зеландии; лесная растительность острова носит почти субтропический характер (и это южнее 45° ю. ш.). Очень мала площадь субантарктических типов растительности, формирующихся в совершенно иных условиях и отличающихся от субарктических типов по набору жизненных форм, ритмике, составу доминантов. Таким образом, на схеме К.Тролля хорошо прослеживаются изменения по широтному градиенту (что отражает изменения в теплообеспеченности) и по градиенту континент -океан, что отражает изменения, связанные с нарастанием континентальности климата. В живом покрове прослеживается и еще один важнейший, высотный градиент, с которым связаны изменения температуры и осадков и соотношения этих показателей при подъеме в горы. В соответствии с обеспеченностью теплом и влагой идут изменения растительности и животного населения с высотой, определяя высотную поясность. Кроме того, вследствие изменения атмосферного давления и, соответственно, парциального давления кислорода и углекислого газа наблюдаются специфические адаптации горных организмов как на уровне физиологических процессов, так и приспособительных адаптации при выработке жизненных форм. В целом для растительного покрова и животного населения в горах характерна смена поясов от подножия горных массивов до их вершины (рис. 24). 132 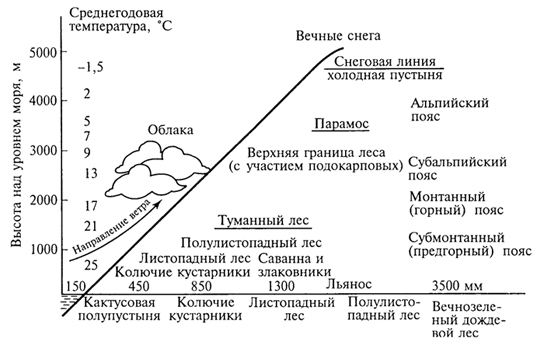 Рис. 24. Высотно-зональное распределение растительного покрова Венесуэлы (G.Walter, S.-W. Вгескlе, 1985) Диапазон условий, в которых формируются биоценозы в горах, огромен. На крутых склонах со скальными породами в мелких трещинах могут существовать лишь немногие виды, слабо осваиваются растительностью курумы, подвижная поверхность которых малодоступна для жизни. В то же время на относительно пологих склонах с развитыми почвами формируется сомкнутый растительный покров. Наиболее суровы условия жизни в верхних поясах гор, особенно выше границы распространения лесной растительности. Биоценозы альпийских и субальпийских лугов, высокогорных тундр, субнивального пояса (по нижней кромке вечных снегов) характеризуются своеобразием видового состава. Здесь распространены собственно высокогорные виды, характеризующиеся специфическими адаптациями (для растений высокогорий употребляется термин "орофиты"). Сообщества высокогорий могут рассматриваться как островные, поскольку зачастую они удалены от сходных с ними сообществ на значительные расстояния. В горах, расположенных в странах с влажным климатом, нередко один и тот же тип растительности характеризует склоны разных экспозиций, например пояса бука или ели в Альпах, пояс даурской лиственницы в горах Восточной Сибири. Асимметричное расположение поясов с выпадением части их на склонах определенных экспозиций характерно для гор стран с сухим или умеренно влажным климатом. Так, в альпийском поясе Заилийского 133 Алатау имеются и альпийские луга, и сообщества арчи, и ивняки. На северном склоне хребтов Тянь-Шаня нередок еловый лес, на южном - степь на тех же высотах. На высоте 3000 - 3500 м на склонах Алайского хребта можно встретить в зависимости от экспозиции арчовники, типчаковые степи, пятна пустынной растительности. Иногда даже трудно дать поясу название по господствующему растительному сообществу. Высотные типы сообществ, сменяющие друг друга по мере подъема (например, в Европе широколиственные листопадные, еловые и елово-пихтовые леса, пояс альпийских лугов), характеризуются значительно меньшей протяженностью по сравнению с широтными зональными типами, их ширина измеряется сотнями метров. Глобальные закономерности изменения набора поясов и положения снеговой линии в горах разных широт представлены на рис. 25. Различия в структуре биоценотического покрова суши хорошо видны и при сравнении зональных спектров на различных материках. Сложная система широтных зон и типов поясности растительного покрова, соответствующая тому или иному материку, в значительной степени обусловлена климатом. Каждый материк отличается своей структурой зональности, что определяется конфигурацией материка, его размерами, характером расположения орографических барьеров и т.д. 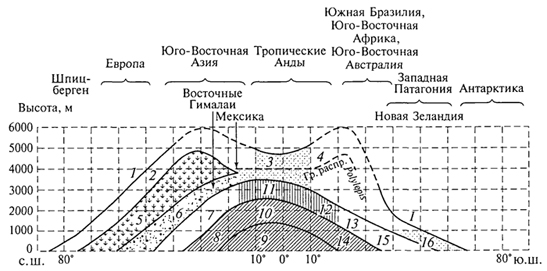 Рис. 25. Схема высотной поясности растительности земного шара от Арктики до Австралии, составленная К.Троллем (по Г.Вальтеру, 1968): 1 - снеговая линия; 2 -тундры, фьельды, альпийская растительность; 3 -парамос; 4 -пуна; 5 -бореальные хвойные леса; 6 -летнезеленые и хвойные леса; 7- летнезеленые и лавровые леса; 8 -субтропические лавровые леса; 9 -тропические дождевые леса; 10- тропические горные леса; 11 -леса пояса облаков; 12- леса из подокарпусов; 13 -леса из араукарий; 14- субтропические дождевые леса; 15- умеренные дождевые леса; 16 -субантарктические леса; отчетливо прослеживается асимметрия высотных поясов 134 Существенное влияние на региональную дифференциацию растительности и животного населения оказывают присущие им особенности циркуляции атмосферы, а также многие биогеографические факторы. В силу этого для различных регионов того или иного материка выделяются свои ряды широтной зональности, и наибольшее число зон характерно для внутриконтинентальных областей. Так, для западной и восточной части Русской равнины выделяются два ряда широтной зональности (от Мурманского побережья до берегов Черного моря и от Печорского побережья до Каспия). Особый ряд широтной зональности свойствен Западно-Сибирской низменности. Местные особенности климатов усложняют во многих регионах проявление зональной дифференциации, приводят к появлению специфических типов сообществ, замещению одних типов другими. В зоне тайги Евразии, например, с запада на восток происходит смена темнохвойных лесов (еловых, пихтовых, кедровых) светлохвойными. Основные массивы темнохвойных лесов сосредоточены в областях умеренно холодного и достаточно влажного климата с ослабленной континентальностью, со сравнительно прохладным летом и высокой влажностью воздуха и почв в вегетационный период. В континентальных районах Южной Сибири темнохвойные леса преимущественно тяготеют к горам (Алтай, Саяны, горы Прибайкалья), где с высотой возрастает влажность воздуха и несколько умеряется континентальность (рис. 26). Область широкого распространения в горах Восточной Сибири и Дальнего Востока своеобразной темнохвойной формации - зарослей кедрового стланика, по всей вероятности, обусловлена проникновением в эти районы на больших высотах воздуха тихоокеанского муссона, что определяет высокую влажность во время вегетационного периода и большое количество осадков зимой (рис. 27). Благодаря утепляющему воздействию снега кедровый стланик с пригнувшимися к земле ветвями благополучно перезимовывает (В.Б.Сочава, 1983). В континентальных районах Сибири, для которых характерны сухость зимнего холодного воздуха, краткий вегетационный период, холодные почвы, часто на небольшой глубине подстилаемые вечной мерзлотой, широкое распространение получают лиственничные леса. Связь лиственничных лесов с резко континентальным климатом показана на рис. 28; видно, что большая часть лиственничных лесов Евразии располагается в области, где коэффициент континентальности превышает 10. В связи с региональными особенностями природных условий происходит смещение границ зон и подзон. В центральной, самой континентальной части Евразии, степи проникают далеко на север, в Якутии они встречаются даже к северу от 60° с.ш. Северная граница средней тайги на Русской равнине и в Западной Сибири 135 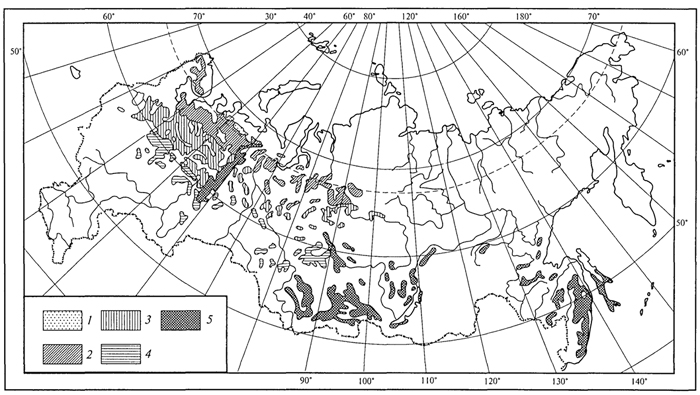 Рис. 26. Распространение темнохвойных лесов в России (В.Б.Сочава, 1956): 1 - предтундровые еловые редколесья; 2 -северотаежные еловые леса; 3 -среднетаежные еловые леса (в Сибири - еловые с участием пихты); 4 -южнотаежные еловые леса (в Сибири - елово-кедрово-пихтовые); 5 - горнотаежные кедровые, пихтовые, еловые леса (на Дальнем Востоке с участием широколиственных пород) 136 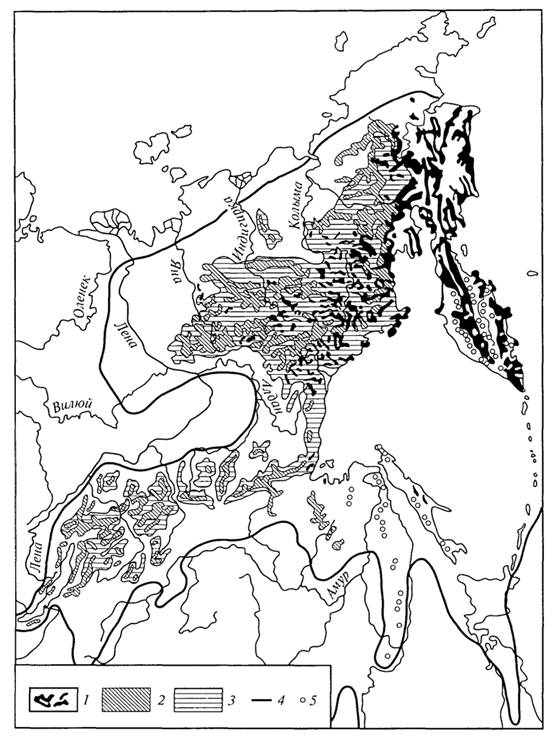 Рис. 27. Районы распространения сообществ кедрового стланика в Восточной Сибири (В.Б.Сочава, 1956): 1 - заросли кедрового стланика; 2 -горные тундры в сочетании с зарослями кедрового стланика; 3 -горные редколесья из лиственницы, местами с подлеском из кедрового стланика; 4 -граница распространения кедрового стланика; 5 -заросли кедрового стланика, встречающиеся на небольших площадях 137 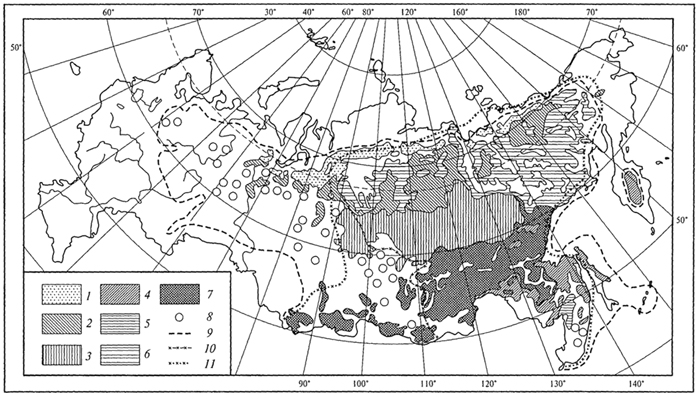 Рис. 28. Распространение лиственничных лесов в Северной Евразии (В.Б.Сочава, 1956): 1 - предтундровые лиственничные редколесья; 2 -северотаежные лиственничные леса; 3- среднетаежные лиственничные леса; 4 -среднетаежные багульниковые и сфагновые лиственничные леса (мари); 5 - южнотаежные лиственничные леса; 6 -горные редкостойные лиственничные леса; 7 -горные таежные лиственничные леса, местами кедрово-лиственничные; 8 -лиственничные леса на небольших площадях; 9 -обобщенная граница распространения всех видов лиственницы; 10 -восточная граница распространения лиственницы сибирской; 11 - изолиния показателя средней континентальности климата (по Н.Н.Иванову) 138 сравнительно хорошо совпадает с южным пределом частого вторжения на материк арктического воздуха. На Западно-Сибирской низменности, где эти вторжения летом обычны и значительны, граница северной тайги заметно смещена к югу. Северная граница южных пустынь тоже совпадает с климатическим рубежом, к югу от которого зимы более умеренные, а приходящие из Ирана циклоны активизируются при приближении арктического фронта, что бывает зимой и ранней весной. На равнинах это благоприятствует развитию эфемерово-полынных пустынь. Не меньшее влияние на особенности структуры живого покрова на региональном уровне оказывают геолого-геоморфологические условия. По отношению к климатическим условиям это связи второго порядка, но вместе с тем они крайне существенны для понимания закономерностей формирования структуры живого покрова. Зональный ряд растительности Западно-Сибирской равнины с типичными для нее водораздельными болотами, заболоченными лесами, займищами, солонцеватыми степями сформировался в условиях компенсированных поднятий и опусканий, которые происходили здесь на протяжении антропогена. На Русской равнине, где в тот же период преобладали положительные движения земной коры, на дренированных водоразделах типичны зеленомошные и мелкотравные темнохвойные леса, дубравы, луговые и разнотравно-дерновиннозлаковые степи. Экологическая среда растительного покрова, а тем самым и его структурные особенности формировались в типичных для региона геолого-геоморфологических условиях, определяющих степень дренированности, глубину залегания грунтовых вод, развитие почвенно-геохимических процессов (В.Б.Сочава, 1983). Огромное влияние на особенности дифференциации растительного покрова оказывает гранулометрический и химический состав почвообразующих пород и почв. Как известно, с гранулометрическим составом связаны водно-физические особенности почв, влияющие и на их тепловые свойства. Поэтому так велики различия в биоценозах, развивающихся, например, на почвах тяжелого (глины, суглинки) и легкого (пески) состава. В пределах одной и той же зоны на различных по составу почвах и почвообразующих 139 породах в качестве доминантов и эдификаторов выступают разные виды, формирующие различные варианты зональных типов сообществ. И ельники-зеленомошники, и сосняки-зеленомошники - типичные биоценозы тайги, однако сосняки более тяготеют к легким по составу почвам, ельники - к более тяжелым. Систематический состав биоценозов зоны включает в себя виды различного географического происхождения и распространения, в соответствии с положением зоны относительно основных центров флоро- и фауногенеза, в связи с чем в разных частях зоны формируются сходные, но не идентичные биоценозы. Так, на протяжении лесостепи Евразии с запада на восток в качестве лесообразователей выступают дуб черешчатый, в Западной Сибири - береза, а еще восточнее - лиственница. При сходстве ценотической организации биоценозов (принадлежности к одним и тем же или близким жизненным формам и контроль макроклимата) различные виды вследствие своих эколого-биологических особенностей формируют и несколько различные по структуре и динамике биоценозы. Таким образом, структура живого покрова зоны характеризуется существенными региональными различиями, обусловленными изменением климатических и геолого-геоморфологических условий, а также особенностями формирования флор и фаун. В отечественной биогеографии существует давняя традиция при анализе структуры биоценотического покрова зон различать зональные, экстразональные и интразональные биоценозы. Зональные биоценозы развиваются на плакорах (термин плакор введен в науку известным русским лесоводом и биогеографом Г. Н. Высоцким) - хорошо дренированных водораздельных равнинах с почвами среднего механического состава - легкосуглинистыми или супесчаными. Иными словами, радиационный режим и условия увлажнения на плакорах типичны для зоны. Экстразональные - это те биоценозы, которые в специфических условиях местообитания "выходят" за пределы своей зоны, чаще всего проникая в смежные. Так, фрагменты широколиственных лесов европейской части России в степной зоне тяготеют к склонам балок и речных долин, где условия увлажнения более благоприятны для них, чем на плакорах, и где комплекс экологических условий в известной степени воссоздает условия смежной зоны. Здесь они являются экстразональными. К интразональным биоценозам относятся такие, которые нигде не выходят на плакоры и не образуют собственной зоны. Интразональные биоценозы связаны в своем распространении с местообитаниями, где условия увлажнения по тем или иным причинам существенно отличаются от таковых на плакорах. Яркий пример таких биоценозов - пойменные (периодически заливаемые) луга, растительность и животное население которых существенно отличаются 140 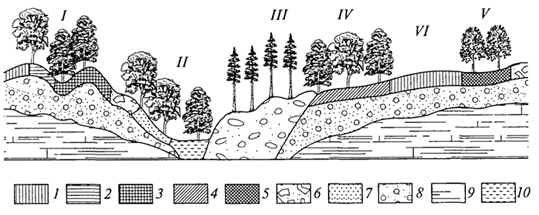 Рис. 29. Закономерности распространения растительности лесостепной зоны в связи с почвенно-геоморфологическими условиями (В.Н.Сукачев, 1975): I- комплексы дубовых лесов на хорошо дренируемых возвышенностях или склонах; II - пойменные и террасные леса с дубом и др.; III - сосновые леса на бедных песчаных почвах, со сфагновыми болотами в переувлажненных низинах; IV -сосново-дубовые леса на суглинистых почвах; V -осиновые рощи в небольших низинах (подах), места скопления талых вод (почвы в центральной части выщелоченные); VI -луговые степи. Почвенно-геоморфологические условия: 1 - мощные слабодренированные черноземы на плакорных пространствах; 2 - хорошо дренируемые деградированные черноземы; 3 - темно-серые лесные почвы; 4 -водопроницаемые песчано-суглинистые лесные почвы; 5 - светлосерые лесные почвы; 6 -солонцы на речных террасах; 7- флювиогляциальные пески; 8-моренные отложения или лёссовидные суглинки; 9 -доледниковые отложения; 10 -аллювий речных долин от зональных. Интразональными являются и биоценозы литоралей: засоленные приморские луга в умеренных широтах, мангровые формации в тропическом поясе. Развитие интразональных биоценозов подчиняется важнейшим географическим закономерностям и, например, между пойменными лугами, расположенными в степной и таежной зонах, существуют известные различия. Еще заметнее различия между биоценозами болот, расположенных в разных зонах. Контрастность между зональными и интразональными биоценозами тем выше, чем ощутимее недостаток влаги в зональных местообитаниях. Неглубокие суффозионные западинки среди южных степей резко выделяются физиономически (яркой зеленью до середины лета, обилием цветущих видов) и по видовому составу. Как правило, в более южных подзонах степей в западинках еще встречаются виды, свойственные северным плакорным местообитаниям. Есть и целые группы видов, связанные преимущественно (или только) с интразональными местообитаниями. На топологическом (ландшафтном) уровне выявляются сочетания сообществ, обусловленные сменой экологических условий по рельефу, возникают территориально сопряженные ряды биоценозов (рис. 29). 141 |