Биология клетки. Размножение организмов
 Скачать 15.49 Mb. Скачать 15.49 Mb.
|

|
7. Типы клеточной организации жизни и происхождение эукариотической клетки, сходство и отличие прокариот и эукариот. Среди всего многообразия ныне существующих на Земле организмов выделяют две группы: вирусы и фаги, не имеющие клеточного строения; все остальные организмы представлены разнообразными клеточными формами жизни. Различают два типа клеточной организации: прокариотический и эукариотический. Клетки прокариотического типа устроены сравнительно просто. В них нет морфологически обособленного ядра, единственная хромосома образована кольцевидной ДНК и находится в цитоплазме; мембранные органеллы отсутствуют (их функцию выполняют различные впячивания плазматической мембраны); в цитоплазме имеются многочисленные мелкие рибосомы; микротрубочки отсутствуют, поэтому цитоплазма неподвижна, а реснички и жгутики имеют особую структуру. Особенности структуры прокариотических клеток определяют специфический характер процессов обмена веществ, жизнедеятельности и размножения. К прокариотам относят бактерий. Они были единственной формой жизни на Земле по крайней мере в течение 2 млрд. лет. Одну из групп фотосинтезирующих бактерий (синезеленые водоросли, или цианобактерии) раньше относили к водорослям. Однако в настоящее время их рассматривают как специфическую группу бактерий. Большинство современных живых организмов относится к одному из трех царств — растений, грибов и животных, объединяемых в надцарство эукариот. Для растительных клеток характерно наличие толстой целлюлозной клеточной стенки, различных пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. В качестве резервного питательного углевода клетки растений запасают крахмал. В клетках грибов клеточная оболочка содержит хитин, в цитоплазме имеется центральная вакуоль, отсутствуют пластиды. Только у некоторых грибов в клеточном центре встречается центриоль. Главным резервным полисахаридом является гликоген. Животные клетки имеют, как правило, тонкую клеточную стенку, не содержат пластид и центральной вакуоли, для клеточного центра характерна центриоль. Запасным углеводом является гликоген. В зависимости от количества клеток, из которых состоят организмы, последние делят на одноклеточные и многоклеточные. Одноклеточные организмы состоят из одной-единственной клетки, выполняющей функции целостного организма. Многие из этих клеток устроены гораздо сложнее, чем клетки многоклеточного организма. Одноклеточными являются все прокариоты, а также простейшие, некоторые зеленые водоросли и грибы. Тело многоклеточных организмов состоит из множества клеток, объединенных в ткани, органы и системы органов. Клетки многоклеточного организма специализированы для выполнения определенной функции и могут существовать вне организма лишь в микросреде, близкой к физиологической (например, в условиях культуры тканей). Клетки в составе многоклеточного организма различаются по размерам, форме, структуре и выполняемым функциям. Несмотря на индивидуальные особенности, все клетки построены по единому плану и имеют много общих черт. 8. Гипотезы происхождения эукариотических клеток Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0—1,4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение. Наиболее популярна в настоящее время симбиотическая гипотезапроисхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот,способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней. 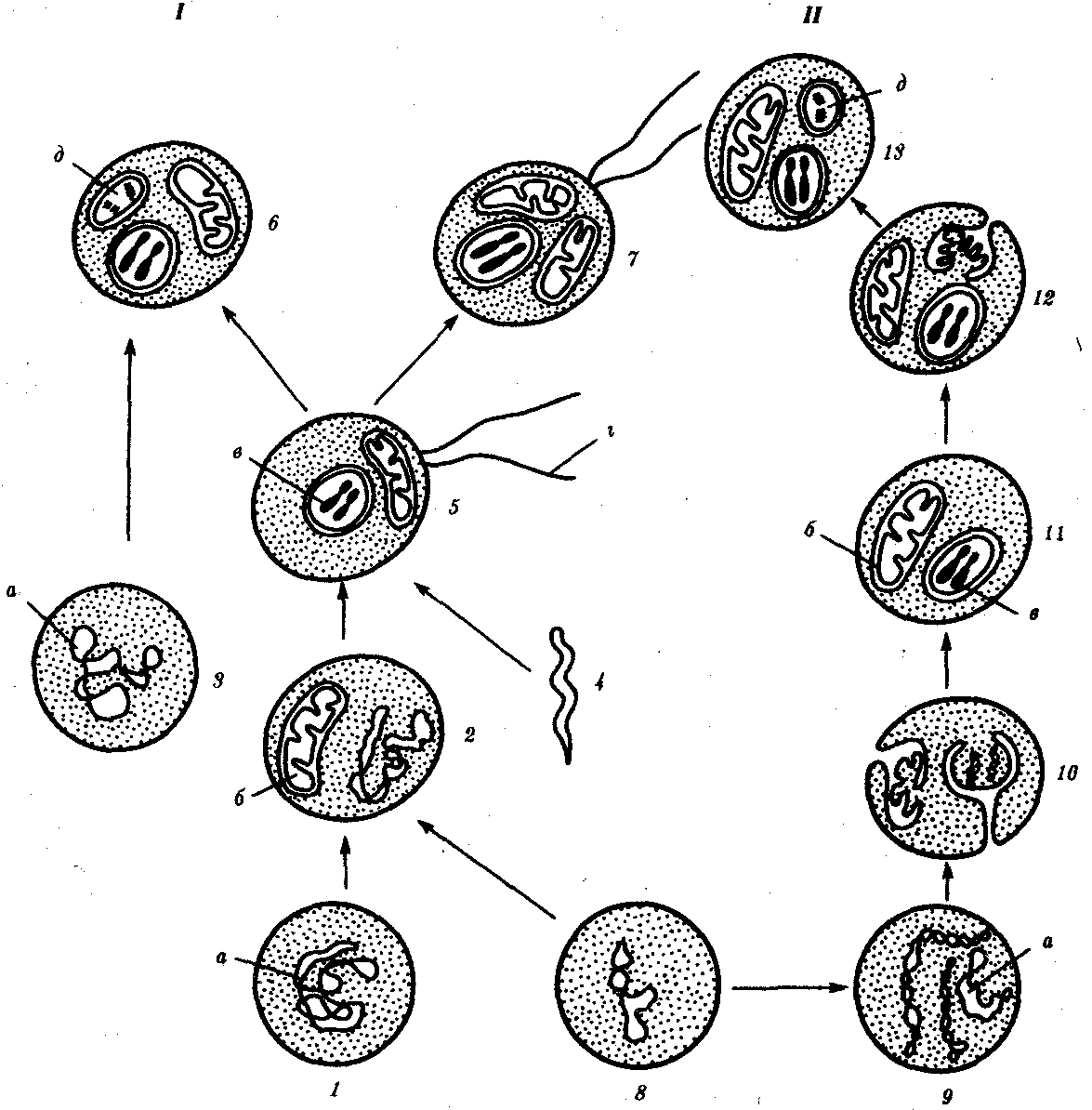 Рис. 1.4. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам: 1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 —сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия),9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 —инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а—ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г —жгутик, д — хлоропласт Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза. Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли. Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0». Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания. Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов). Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4).Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки. История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100-1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости. При переходе к эукариотическому типу усложняется механизм регуляции жизнедеятельности клетки, что на уровне генетического материала проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками. В итоге стало возможным считывать биологическую информацию по частям с разных групп генов в разном их сочетании в различных типах клеток и в разное время. В бактериальной клетке, напротив, одновременно считывается до 80—100% информации генома. В клетках взрослого человека в разных его органах транскрибируется от 8—10% (печень, почка) до 44% (головной мозг) информации. Использованию биологической информации частями принадлежит исключительная роль в эволюции многоклеточных организмов, так как именно это позволяет разным группам клеток специализироваться по различным функциональным направлениям. Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов. Среди цитофизиологических особенностей эукариот, увеличивающих их эволюционные возможности, необходимо назватьаэробное дыхание, которое также послужило предпосылкой для развития многоклеточных форм. Интересно, что сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). Названная концентрация является необходимым условием аэробного дыхания. В условиях усложнения генетического аппарата эукариот, увеличения суммарного количества ДНК и распределения ее по хромосомам трудно переоценить значение возникновения в эволюции митоза как механизма воспроизведения в поколениях генетически сходных клеток. Появление вследствие эволюционных преобразований митоза такого способа деления клеток, как мейоз, дающего возможность сохранить постоянство хромосом в ряду поколений, наилучшим образом решило проблему размножения многоклеточных организмов. Связанный с мейозом переход к половому размножению усилил эволюционную роль комбинативной изменчивости, способствовал увеличению скорости эволюции. Благодаря отмеченным особенностям за 1 млрд. лет эволюции эукариотический тип клеточной организации дал широкое разнообразие живых форм от одноклеточных простейших до млекопитающих и человека. 9. Современные доказательства симбиотического происхождения эукариот 10. Сущность клеточной теории, современное состояние клеточной теории и её значение для биологии. Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений, животных и остальных живых организмов с клеточным строением, в котором клетка рассматривается в качестве общего структурного элемента живых организмов. Клеточная теория
Шлейден предложил считать ядро наиболее постоянной структурой клетки. Многие положения оказались неверными (положение 3). Клетка стала изучаться. Клеточная теория оказала большое влияние на биологию и медицину. 1858 – Рудольф Вирхов опубликовал свой труд. Если существует живая клетка, то она произошла от клетки. « Каждая клетка от клетки». Применил свои положения теории в клеточной патологии. Современная клеточная теория
Активность организма зависит от клетки, в клетке хранится, перерабатывается наследственная информация. Клетка – основная единица, через которую проходит, запасается и перерабатывается энергия, вещество. Клетка простейшего практически бессмертна. При половом размножении вечны гаметы. В клетке клеточные структуры связаны между собой, все биохимические процессы происходят в соответствующей структуре. Современная клеточная теория исходит из того, что клеточная структура является главнейшей формой существования жизни, присущей всем живым организмам, кроме вирусов. Совершенствование клеточной структуры явилось главным направлением эволюционного развития как у растений, так и у животных, и клеточное строение прочно удержалось у большинства современных организмов. Вместе с тем должны быть подвергнуты переоценке догматические и методологически неправильные положения клеточной теории: Клеточная структура является главной, но не единственной формой существования жизни. Неклеточными формами жизни можно считать вирусы. Правда, признаки живого (обмен веществ, способность к размножению и т.п.) они проявляют только внутри клеток, вне клеток вирус является сложным химическим веществом. По мнению большинства учёных, в своём происхождении вирусы связаны с клеткой, являются частью её генетического материала, "одичавшими" генами. Выяснилось, что существует два типа клеток - прокариотические (клетки бактерий и архебактерий), не имеющие отграниченного мембранами ядра, и эукариотические (клетки растений, животных, грибов и протистов), имеющие ядро, окружённое двойной мембраной с ядерными порами. Между клетками прокариот и эукариот существует и множество иных различий. У большинства прокариот нет внутренних мембранных органоидов, а у большинства эукариот есть митохондрии и хлоропласты. В соответствии с теорией симбиогенеза, эти полуавтономные органоиды - потомки бактериальных клеток. Таким образом, эукариотическая клетка - система более высокого уровня организации, она не может считаться целиком гомологичной клетке бактерии (клетка бактерии гомологична одной митохондрии клетки человека). Гомология всех клеток, таким образом, свелась к наличию у них замкнутой наружной мембраны из двойного слоя фосфолипидов (у архебактерий она имеет иной химический состав, чем у остальных групп организмов), рибосом и хромосом - наследственного материала в виде молекул ДНК, образующих комплекс с белками. Это, конечно, не отменяет общего происхождения всех клеток, которое подтверждается общностью их химического состава. Клеточная теория рассматривала организм как сумму клеток, а жизнепроявления организма растворяла в сумме жизнепроявлений составляющих его клеток. Этим игнорировалась целостность организма, закономерности целого подменялись суммой частей. Считая клетку всеобщим структурным элементом, клеточная теория рассматривала как вполне гомологичные структуры тканевые клетки и гаметы, протистов и бластомеры. Применимость понятия клетки к протистам является дискуссионным вопросом клеточного учения в том смысле, что многие сложно устроенные многоядерные клетки протистов могут рассматриваться как надклеточные структуры. В тканевых клетках, половых клетках, протистах проявляется общая клеточная организация, выражающаяся в морфологическом выделении кариоплазмы в виде ядра, однако эти структуры нельзя считать качественно равноценными, вынося за пределы понятия «клетка» все их специфические особенности. В частности, гаметы животных или растений - это не просто клетки многоклеточного организма, а особое гаплоидное поколение их жизненного цикла, обладающее генетическими, морфологическими, а иногда и экологическими особенностями и подверженное независимому действию естественного отбора. В то же время практически все эукариотические клетки, несомненно, имеют общее происхождение и набор гомологичных структур - элементы цитоскелета, рибосомы эукариотического типа и др. Догматическая клеточная теория игнорировала специфичность неклеточных структур в организме или даже признавала их, как это делал Вирхов, неживыми. В действительности, в организме кроме клеток есть многоядерные надклеточные структуры (синцитии, симпласты) и безъядерное межклеточное вещество, обладающее способностью к метаболизму и потому живое. Установить специфичность их жизнепроявлений и значение для организма является задачей современной цитологии. В то же время и многоядерные структуры, и внеклеточное вещество появляются только из клеток. Синцитии и симпласты многоклеточных - продукт слияния исходных клеток, а внеклеточное вещество - продукт их секреции, т.е. образуется оно в результате метаболизма клеток. Проблема части и целого разрешалась ортодоксальной клеточной теорией метафизически: всё внимание переносилось на части организма — клетки или «элементарные организмы». Целостность организма есть результат естественных, материальных взаимосвязей, вполне доступных исследованию и раскрытию. Клетки многоклеточного организма не являются индивидуумами, способными существовать самостоятельно (так называемые культуры клеток вне организма представляют собой искусственно создаваемые биологические системы). К самостоятельному существованию способны, как правило, лишь те клетки многоклеточных, которые дают начало новым особям (гаметы, зиготы или споры) и могут рассматриваться как отдельные организмы. Клетка не может быть оторвана от окружающей среды (как, впрочем, и любые живые системы). Сосредоточение всего внимания на отдельных клетках неизбежно приводит к унификации и механистическому пониманию организма как суммы частей. Очищенная от механицизма и дополненная новыми данными клеточная теория остается одним из важнейших биологических обобщений. 11. Биологическая мембрана, молекулярные организация и функции. Транспорт веществ через мембрану. Модели транспорта. Биомембраны отделяют клеточное содержимое от внешней среды, регулируют обмен между клеткой и средой, делят клетку на отсеки или компартменты, предназначенные для раздельного протекания тех или иных метаболических путей. Ряд химических реакций в клетке протекает непосредственно на мембранах. На них располагаются и рецепторы для распознавания внешних сигналов из окружающей среды. Строение мембран. Существует несколько моделей строения мембран. В настоящее время принята жидко-мозаичная модель Сингера и Николсона. Согласно этой модели, липиды образуют бислой, в котором плавают белки. Из-за различной степени погружения белковых молекул образуется своеобразная мозаика, что и отражено в названии модели. Если суммировать известные нам данные о строении биологических мембран, можно выделить следующие положения: 1.разные типы мембран отличаются по своей толщине, но в большинстве случаев она составляет 5-10 нм. 2.мембраны - это липопротеиновые структуры, на внешней стороне к липидам и / или белкам прикреплены углеводные компоненты, на долю которых приходится от 2 до 10%. 3.липиды спонтанно образуют бислой, так как их молекулы имеют полярные головы и неполярные хвосты. 4.мембранные белки выполняют разнообразные функции. 5.углеводные компоненты образуют надмембранный комплекс, называемый гликокаликсом, участвуют в механизмах распознования. Состав мембранных липидов достаточно разнообразен. Мембранные липиды обеспечивают "жидкое", "рыхлое" состояние бислоя. Жидкое состояние мембран -важный фактор, обеспечивающий транспорт, активность мембраносвязанных белков и процесс слияния отдельных мембран друг с другом. Липидный бислой обладает рядом важных свойств. К их числу относятся: способность к самосборке, полупроницаемость и диэлектрические свойства. Способность к самосборке позволяет быстро восстанавливать поврежденную целостность клеточной мембраны, полупроницаемость лежит в основе транспорта через мембраны. Диэлектрические свойства липидов не позволяют передавать заряд, поэтому на наружной и внутренней поверхности мембраны может образовываться разность потенциалов, которая называется мембранным потенциалом. Мембранные белки условно можно разделить на две группы. К первой относятся структурные белки, ко - второй - белки, выполняющие наряду со структурными и другие функции. В их числе следует особо выделить транспортную функцию, функцию маркировки и рецепции, ферментативную функцию, а также участие в преобразовании энергии в процессах дыхания и фотосинтеза. Большую роль играют комплексы белков с углеводами, называемые гликопротеинами. На свободных поверхностях гликопротеинов, ориентированных на внешнюю сторону мембраны, находятся олигосахаридные цепи, напоминающие антены. Их функция – распознавание внешних сигналов, т.е. они играют роль информационных молекул. С функцией распознавания связан процесс регуляции иммунного ответа, объединение клеток в ткани. Кроме гликопротеидов функцию распознавания выполняют и гликолипиды. Нарушение гликокаликса приводит к внедрению в клетку вирусов и т.д. Одной из наиболее важной функцией мембран является транспортная. Известны 4 процесса транспорта через плазматическую мембрану, причем два из них не требуют затрат энергии и называются пассивными, а два последние - активные, т.е. протекают с потреблением энергии. Простейшим видом транспорта через мембрану является осмос или диффузия воды. Другим видом пассивного транспорта является диффузия по градиенту концентрации, т.е. из области с высокой концентрацией в более низкую. Этим видом транспорта в клетку попадают малые молекулы (с массой не более 150 Да), а также кислород, углекислый газ и азот. Эти вещества проходят по туннельным мембранным белкам. У таких молекул имеется канал, пропускающий вещества определенного типа. Пассивная диффузия характерна для молекул или ионов, не имеющих заряда. Другим видом пассивной диффузии является растворение вещества в липидном бислое. Так в клетку попадают спирты, глицерол. Частным случаем является облегченная диффузия. В этом случае веществу помогает пройти мембранный барьер специфический транспортный белок или белок - переносчик. Этот белок соединяется на одной стороне мембраны с веществом, а на другой стороне - его отдает. Такой транспорт веществ осуществляется по электрохимическому градиенту. Примером является транспорт глюкозы в эритроциты. Принцип работы белков- переносчиков. Это образование канала в молекуле белка из-за конформационных подвижек, или механизм понг-пинга. В стадии понг - участок для соединения белка - переносчика с веществом открыт во внешнюю среду, пинг – участок закрывается из-за конформационных изменений белка и открывается вовнутрь клетки. Активный транспорт - это сопряженный с потреблением энергии процесс переноса веществ против электрохимического градиента. Например, рассмотрим ионы натрия, калия и хлора. Их содержание в плазме крови намного отличается от такового в эритроцитах. Исследования последних лет показали, что в клетке активно действует натриевый насос, выкачивающий ионы натрия из клеток. Чаще всего он сопряжен с калиевым насосом, который закачивает ионы калия вовнутрь клетки. Такой объединенный насос называется калий, натриевый насос. Насос - это белок, пронизывающий мембрану насквозь. С внутренней стороны мембраны к нему подходят ионы натрия и АТФ, снаружи - ионы калия. Считается, что обмен ионами происходит путем конформационных изменений белка. Известны также и другие виды белков-насосов. Механизм работы транспортных белков. Различают 3 вида механизма работы транспортных белков: унипорт, симпорт и антипорт. При унипорте белок переносит вещество через мембрану, при симпорте перенос вещества или иона сопряжен с переносом другого иона. Примером симпорта является перенос глюкозы в клетку, сопряженный с переносом натрия. При антипорте происходит одновременный перенос как вовнутрь, так и из клетки. Примером является натрий, калиевый насос. Принцип работы транспортных белков -обычно понг-пинг. Через плазматическую мембрану транспортируются и макромолекулы. Процесс, с помощью которого клетки захватывают крупные молекулы, называется эндоцитозом. Процесс выхода крупных молекул из клетки называется экзоцитозом. Общим для этих видов транспорта является то, транспортируемое вещество окружено плазматической мембраной и находится в виде пузырька или везикулы. Механизм образования везикулы и её судьба в клетке зависит от типа эндоцитоза. Эндоцитоз. Эндоцитоз можно разделить на 2 основных типа: фагоцитоз и пиноцитоз. Пиноцитоз присущ всем клеткам. С его помощью клетка поглощает жидкости и мелкие гранулы. При фагоцитозе происходит поглощение крупных частиц:вирусов, бактерий, клеток или их обломков. Фагоцитоз осуществляется с участием специализированных клеток: макрофагов и гранулоцитов. Механизм эндоцитоза: при захвате вещества из межклеточного пространства происходит впячивание или инвагинация плазматической мембраны, образуется эндоцитозная везикула, похожая на колбу. Шейка везикулы сливается, отшнуровывается от мембраны. Судьба везикул различна: они могут направляться к комплексу Гольджи или транспортируются к лизосомам, сливаются с ними, образуя вторичные лизосомы или фаголизосомы. Эндоцитоз подразделяется 2 типа: жидкофазный неспецифичный и адсорбционный рецепторный (с очень быстрым избирательным захватом макромолекул). Название и судьба везикул, образующихся при адсорбционном эндоцитозе, зависят от типа поглощаемого вещества. Если поглощаемым веществом является гормон, то такая везикула снабжена рецепторами, позволяющим ей достичь конкретного места в клетке, например, комплекса Гольджи. Такая везикула называется рецептосомой. Если содержимое везикулы сливается с лизосомой, то такие везикулы называются окаймленными пузырьками, так как они содержат белок - клатрин. Опосредуемый рецепторами эндопитоз имеет и свои теневые стороны, поскольку ряд вирусов, например, СПИДа, гепатита, полимиелита атакуют клетки именно по этому механизму. Экзоцитоз имеет большое значение для клетки. С его помощью клетка, например, обновляет свои мембраны, осуществляет секреторную деятельность. Механизм экзоцитоза: вещества в везикулах отпочковываются от комплекса Гольджи или от эндоплазматического ретикулума, транспортируются к мембране, сливаются с ней, после чего содержимое везикулы выполняет свое предназначение. Экзоцитоз может быть непрерывным, так и прерывистым. Вещества, высвобождаемые в процессе экзоцитоза, можно разделить на 3 группы: 1.вещества, связывающиеся с клеточной поверхностью, например, антигены; 2. вещества внеклеточного матрикса, 3.сигнальные молекулы (гормоны, медиаторы) Изменение свойств биомембран, возникающие под влиянием факторов внешней и внутренней среды, лежат в основе развития многих патологических состояний и процессов. Нарушение функции биомембран может быть следствием развития патологических процессов. Это привело к возникновению такого понятия как "патология биомембран" Патология биомембран может быть связана с мембранными липидами, мембранными белками, включая рецепторы, а также с гликокаликсом. Например, биомембраны являются мишенями действия ядов, токсинов, радиоактивного излучения, ультрафиолета. Нарушение функций мембранных рецепторов часто выступает причиной патологии иммунной системы. Механизмы возникновения и развития таких заболеваний как гипертония, атеросклероз и т.д, связаны с нарушением структуры и функции мембранных белков и липидов. Изучение строения, свойств и функций биомембран в условиях нормы позволит лучше усвоить причины и механизмы их нарушения в при действии негативных факторов. |