Бизюк основы нейропсихологии. Бизюк - основы нейропсихологии. Бизюк А. П. Основы нейропсихологии
 Скачать 2.73 Mb. Скачать 2.73 Mb.
|

|
ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ оболочка — неврилемма — вместе с миелиновой окружает лишь волокна периферической части НС. Она имеет клеточное строение и по отношению к поврежденному нервному стволу выполняет регенеративную функцию. Отсутствие неврилеммы у волокон нервных клеток головного и спинного мозга не позволяет поврежденным участкам этих органов морфологически восстанавливаться (рис. 4). По функциональному назначению нейроны делятся на афферентные (сенсорные или рецепторные) — получающие информацию из внешней или внутренней среды, эфферентные (моторные) — посылающие импульсы к исполнительным органам и вставочные — обеспечивающие связь одного нейрона с другим. Помимо нейронов с их отростками, масса головного и спинного мозга приблизительно на 40% состоит из клеточной глиальной ткани, равномерно распределенной по всему их объему и заполняющей пространство между нейронами. Функция глии или нейроглии — кроме опорной — обеспечение трофических функций мозга и обслуживание реактивных свойств нервной ткани — образование рубцов после травм, участие в реакциях воспаления, в формировании опухолей и т. д. Функционирование мозга обеспечивается совместной работой всех клеток, взаимодействие между которыми осуществляется посредством синаптической связи. Синапс — специализированный контакт между нервными клетками, служащий для передачи и преобразования сигналов (информации). Межнейронные синапсы в большинстве случаев образованы окончаниями аксонов одних нервных клеток и телом, дендритами (обычно) или аксонами других. В зависимости от назначения клетки, число ее синаптических связей может достигать 20 000. Передача импульса по нервному волокну, а также его переход от одного нейрона к другому сопровождается сложными химическими и электрохимическими преобразованиями как в самой клетке, так и в синаптических аппаратах. При достижении импульсом окончания аксона оттуда в синаптичес-кую щель выделяется специальное вещество — медиатор (посредник), который связывается со специфическими рецепторами на поверхности следующей нервной клетки и изменяет ее способность генерировать импульсы. Почти к каждому нейрону приходят входы от сотен или тысяч окончаний, причем одни из них оказывают на нейрон возбудительное, а другие тормозное воздействие (повышают порог чувствительности к возбуждающим веществам). В каждый данный момент одни входы активны, а другие бездействуют, и от суммы возбудительных и тормозных эффектов зависит активность очередного нейрона в их цепи или сети. Таким образом каждый нейрон непрерывно оценивает все сигналы, приходящие к нему от других клеток, и выражает результаты этой оценки частотой собственных сигналов. 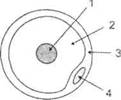 Рис. 4. Схема сечения аксона: / — аксонное волокно; 2— миелиновая оболочка; 3 — неврилемма; 4 — клеточное ядро ОСНОВЫ НЕЙРОПСИХОЛОГИИ 2.2. ОСНОВНЫЕ АНАТОМИЧЕСКИЕ СТРУКТУРЫ МОЗГА В созревшем состоянии головной и спинной мозг, а также весь аппарат периферических нервов с рецепторными органами представляют собой целостную систему, которая анатомически и функционально делится на большое число звеньев. Центральная нервная система (ЦНС) включает в себя спинной и головной мозг. Спинной мозг является цилиндрическим образованием с толстыми стенками, которое на поперечном разрезе представлено двумя типами нервной ткани — серым веществом тел нейронов, сконцентрированных в центральной части в виде буквы «Н» или «бабочки», и белым веществом, образованным пучками восходящих и нисходящих аксонов и дендритами. «Крылья бабочки» разделены на два передних и два задних рога. Передние рога содержат тела моторных нейронов, эфферентные аксоны которых направляются в составе спинномозговых нервов к мышцам. Задние рога содержат клетки промежуточных нейронов, к которым подводятся афферентные волокна, доставляю-. щие чувствительные импульсы с периферии. Топография белого вещества на поперечных срезах спинного мозга такова, что позволяет говорить о передних, задних и боковых столбах, расположенных в соответствующих промежутках между передними и задними рогами (рис. 5). В срединных отделах серого вещества проходит центральный канал, содержащий спинномозговую жидкость. Анализ организации нейронных цепей в задних рогах спинного мозга позволил сделать вывод, что эта часть центральной нервной системы не является только передаточным механизмом, в котором афферентные сигналы транслируются по восходящим путям. Здесь отдельные сенсорные модальности подвергаются сложной переработке, в его многочисленных нейронных цепях и синаптических контактах происходят интегративные процессы, приводящие к тому, что значительное число импульсов, входящих в спинной мозг, отфильтровывается и не достигает больших полушарий. Головной мозг (рис. 6 и 7) анатомически ограничен костными структурами черепа и структурно делится на задний, средний, промежуточный и передний. Задний мозг состоит из продолговатого мозга, мозжечка и моста (варолиева) (рис. 8 и 9). Продолговатый мозг анатомически является продолжением спинного, хотя по внутренней структуре и соотношению серого и белого 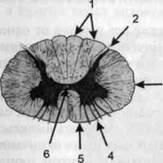 Рис. 5. Горизонтальный разрез верхнего отдела спинного мозга: / — нежный и клиновидный канатики, образующие задний столб; 2 — задний рог; 3 — боковой столб; 4 — передний рог; 5— передний столб; 6 — центральный канал ГЛАВА 2. AHATOMO-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ 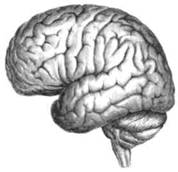 Рис. 6. Общий вид головного мозга со стороны левого полушария 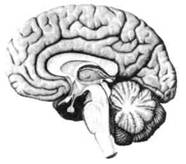 Рис. 7. Общий вид анатомических структур головного мозга на срезе попродольной щели веществ заметно от него отличается. Продолговатый мозг включает в себя ряд ядер, обслуживающих управление дыханием, сердечным ритмом, сосудистым тонусом и рядом других функций, в том числе артикуляторных (поддерживающих согласованную работу органов речи для обеспечения членораздельного произношения). В продолговатом мозгу находится ряд рефлекторных центров, связанных с процессами пищеварения. Это группа центров моторных рефлексов (жева- 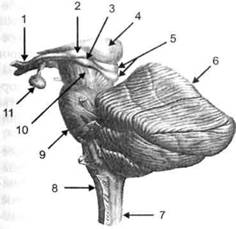 Рис. 8. Средне-нижние отделы головного мозга: J— зрительная хиазма; 2 — латеральное коленчатое тело; 3 — медиальное коленчатое тело; 4 — подушка таламуса (фрагмент); 5 — бугры четверохолмия; 6— мозжечок; 7— спинной мозг; 8 — продолговатый мозг; 9— варолиев мост; 10— ножки мозга; // — гипофиз 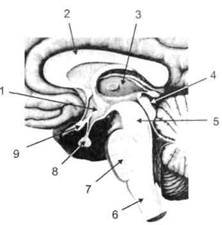 Рис. 9. Средне-нижние отделы головного мозга (срез): / — гипоталамус; 2 — мозолистое тело; 3 -таламус; 4— бугры четверохолмия; 5-ножки мозга; 6— продолговатый мозг; 7-варолиев мост; 8— гипофиз; 9— зрительная хиазма ОСНОВЫ НЕЙРОПСИХОЛОГИИ 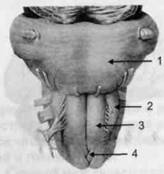 Рис. 10. Передняя поверхность заднего мозга: / — варолиев мост; 2 — оливы; 3— пирамиды; 4— перекрестье пирамид ния, глотания, движении желудка и части кишечника), а также секреторных (слюноотделение, выделение пищеварительных соков желудка, поджелудочной железы и др.). Кроме того, здесь находятся центры некоторых защитных рефлексов: чихания, кашля, мигания, слезоотделения, рвоты. Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц — участвует в осуществлении рефлексов поддержания и восстановления позы тела, так называемых установочных рефлексов. Как филогенетически древнейший отдел головного мозга, он имеет важное значение в осуществлении функций сна. На передней поверхности продолговатого мозга имеются два вертикальных валика, получившие название пирамид. По бокам от них расположена другая пара валиков, обозначаемых как оливы (рис. 10). Мозжечок — двухполушарный орган кзади сверху от продолговатого мозга, имеющий дольчатую структуру и принимающий участие в координации движений, сохранении позы, тонуса и равновесия тела, то есть тех компонентов жизнедеятельности, которые имеют отношение к гравитационным условиям обитания организма. Помимо этого, мозжечок вместе с лобной корой больших полушарий соучаствует в формировании программ движений на основе обратной афферентации, поступающей в него от проприорецепторов (мышечно-суставных), а также вестибулярных, зрительных и тактильных анализаторов. Он же осуществляет и контроль точности движений артикулятор-ного аппарата, косвенно влияя на речевые функции. Имеются свидетельства о роли мозжечка в программировании искусственной координации двух движений. Исследования последних десятилетий позволили выдвинуть предположение, что мозжечок оказывает влияние и на когнитивные процессы. Он осуществляет контроль за «развертыванием» во времени и точным взаимодействием (или координацией) когнитивных операций, то есть обеспечивает быстрое, плавное выполнение психических действий подобно тому, как это происходит в двигательных процессах. Мозжечок включается в решение любого типа задач, где требуются быстрые и последовательные изменения, а также регулирует психическую активность при переходе с одних задач на другие (Ю. В. Зуева, Н. К. Корсакова, Л. А. Калашникова). Варолиев мост — это крупное поперечно-волокнистое образование, охватывающее передне-верхнюю часть продолговатого мозга. В функцию моста входит проведение координирующих импульсов от одного полушария мозжечка к другому для обеспечения согласованных движений мышц на обеих сторонах тела и связь коры больших полушарий с самим мозжечком. В варо- ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ 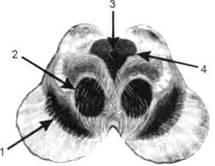 Рис. 11. Горизонтальный срез ножек мозга: /— черная субстанция; 2— красное ядро; 3 — сильвиев водопровод; 4 — центральное серое вещество лиевом мосту поперечно проходят волокна, имеющие отношения к слуховой системе (трапецевидное тело), а продольно — к двигательной. Средний мозг (мезенцефальный), расположенный над мостом, включает в себя ножки мозга (две ветви разделившегося пополам продолговатого мозга) и четверохолмие — четыре округлых выступа на задней поверхности среднего мозга. На ранних этапах эволюции они являлись древними центрами зрения (верхние бугры) и слуха (нижние бугры) (рис. 8 и 9). Средний мозг играет важную роль в регуляции движений глаз, для чего служат несколько ядер, осуществляющих поворот глаз в любом направлении, аккомодацию глаз, фиксацию взгляда на близких предметах путем сведения зрительных осей, реализующих зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету). В каждой из ножек мозга, помимо проводящих волокон, располагаются имеющие отношение к регуляции движений богатая пигментом черная субстанция (регулирует тонус мышц, особенно при выполнении мелких движений пальцами рук) и красное ядро (поддерживает тонус скелетной мускулатуры) (рис. 11). В глубине среднего мозга вдоль задней его поверхности проходит узкий канал, соединяющий 3-й и 4-й желудочки мозга, — сильвиев водопровод. Все отделы между спинным мозгом и промежуточным мозгом (включительно) образуют ствол мозга, в массе которого расположена ретикулярная (се-тевидная) формация, волокна которой переплетаются со всеми проходящими через него афферентными и двигательными путями. Она оказывает энергорегулирующее воздействие на кору больших полушарий и контролирует рефлекторную деятельность спинного мозга (рис. 12). Промежуточный мозг (диэнцефальный) включает в себя два отдела — зрительный бугор (таламус) и подбугорную область (гипоталамус) (рис. 9). Таламус представляет собой сложный парный комплекс серого вещества, образующего пространственную конструкцию, похожую на два сросшихся яйца, каждое из которых составляет единое це- 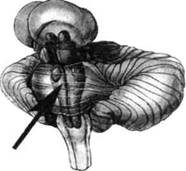 Рис. 12. Ретикулярная формация среднего и заднего мозга (по Ф. Блу- му, А. Лейзерсону, Л. Хофстедтеру) ОСНОВЫ НЕЙРОПСИХОЛОГИИ лое со своим полушарием. Задняя расширяющаяся часть каждой из половин получила названия подушки таламуса. В нижне-боковой ее части имеются два выпячивания — латеральное (наружное) и медиальное (внутреннее) коленчатые тела (ЛКТ и МКТ) (рис. 8). Таламус выполняет функцию коллектора и коммутатора всех возбуждений, поступающих от рецепторов в головной мозг (кроме обоняния), то есть производит предварительный анализ и синтез импульсов от всех органов чувств и через синаптические связи направляет их в соответствующие зоны мозга. Передние отделы зрительного бугра посылают волокна к различным зонам лобных долей, а задние — преимущественно к участкам теменной, а также затылочной области. В составе зрительного бугра принято выделять: (1) два типа специфических ядер — релейные, в которых по пути в кору переключаются сенсорные импульсы, и ассоциативные (интегрирующие, объединительные), в которые информация поступает не с периферии, а после предварительной обработки в других ядрах таламуса; (2) неспецифические, эволюционно более древние, оказывающие модулирующее (изменяющее параметры) влияние на кору. Они входят в систему ретикулярной формации и выступают в роли посредника между стволом мозга и более высокорасположенными структурами, объединяя их в единый функциональный комплекс. Гипоталамус представляет относительно узкий слой мозговой ткани, расположенный в промежутке между зрительным бугром и разрывом ножек мозга. В нем расположены многочисленные высокодифференцированные ядра, регулирующие температуру тела, аппетит, водный баланс, углеводный и жировой обмен, сосудистый тонус и другие вегетативные функции, связанные с обменом веществ, в том числе с нейро-гуморально-гормональным контролем. Здесь же находятся центры, осуществляющие регуляцию сна, сексуального и эмоционального поведения. Гипоталамус играет важнейшую роль в регуляции гомеостаза (постоянства внутренней среды организма), но, помимо решения физиологических задач, проявляет себя и в качестве: 1) главного «сенсорного входа» в эмоциональную систему сигналов из внутренней среды организма; 2) подкоркового интегратора информации, имеющей отношение к реализации эмоций; 3) «выхода» для импульсов, обеспечивающих внешнее выражение эмоциональных состояний. Именно гипоталамус передает во фронтальную кору возбуждение мотива-ционного характера. Исключительно велика роль связи ретикулярной формации с находящимися в гипоталамусе центрами, обеспечивающими пластичность и динамическую устойчивость внутренней среды организма. Кроме того, к структурам гипоталамуса анатомически относят гипофиз — железу внутренней секреции и зрительную хиазму — место неполного перекрестья зрительных нервов (рис. 8 и 9). Передний мозг состоит из двух полушарий, покрытых серым веществом — корой (ее толщина у взрослого человека колеблется от 1 до 5 мм). При рас- -1 П ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ 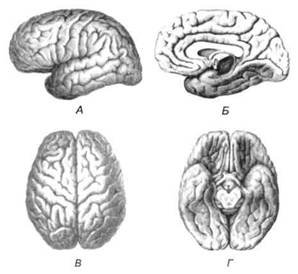 Рис. 13. Латеральная (А), медиальная (Б), конвекситальная (В) и базальная (Г) поверхности больших полушарий смотрении поверхности мозга и для локализации отдельных участков в нейропсихологии принято пользоваться специальной терминологией, обозначающей плоскость или сектор обзора, на которых фиксируется внимание. Кон-векситальный (букв. — выпуклый, здесь — наружный) мозг — поверхность полушарий, видная при-взгляде сверху; базальный мозг — поверхность и анатомические структуры, видные при взгляде снизу; латеральный мозг — поверхности, видные при взгляде сбоку; медиальный мозг— поверхности полушарий, обращенные внутрь продольной щели мозга (рис. 13). В глубине каждого полушария расположены проводящие волокна и подкорковые ядра (базальные ганглии), главные из которых — это полосатое тело (striatum), включающее в себя хвостатое ядро, скорлупу и бледный шар (pallidum). Они имеют достаточно сложную внутримозго-вую архитектуру и объединяются под общим названием — стриопал-лидарной системы (рис. 14). Эта система имеет довольно сложный филогенез, в результате которого из целостного подкоркового образования возникают упомянутые анатомические структуры, причем палли-дум из них является самым древним 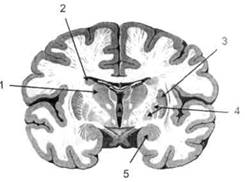 71 Рис. 14. Основные подкорковые ядра на вертикальном срезе больших полушарий говного мозга: / — таламус; 2 — хвостатое ядро; 3 — скорлупа; 4 — бледный шар; 5 — миндалевидное тело ОСНОВЫ НЕЙРОПСИХОЛОГИИ 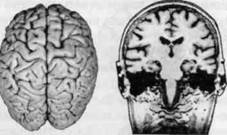 и достаточно примитивным органом (из-за этого его иногда называют па-леостриатумом), а полосатое тело, микроскопическая структура которого приближается к структуре мозговой коры, — молодым (неостриатумом). Поверхность полушарий покрыта бороздами и извилинами, причем величина и форма борозд подвержены значительным индивидуальным колебаниям, вследствие чего не только мозг различных людей, ной полушария одного и того же индивида по рисунку борозд могут заметно отличаться друг от друга (рис. 15). Глубокими постоянными бороздами пользуются для разделения каждого полушария на большие участки, называемые долями. Такими бороздами являются боковая (сильвиева) борозда, центральная (роландова) борозда и те-менно-затылочная борозда, отчетливо обозначенная лишь на поверхности продольной шели мозга, делящей его на два полушария (рис. 16). Основные извилины показаны на рис. 17. Рис. 15. Различия между профилем борозд правого и левого полушарий: слева — препарат мозга, справа — по материалам электронного исследования мозга (вертикальный срез) 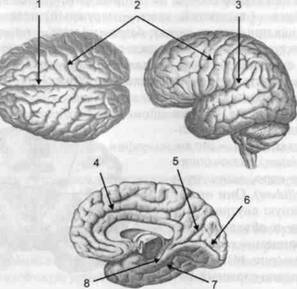 Рис. 16. Основные борозды больших полушарий головного мозга: / — продольная щель; 2— роландова (центральная) борозда; 3— сильвиева (боковая) борозда; 4— поясная борозда; 5— теменно-затылочная борозда; 6— шпорная борозда; 7— коллатеральная борозда; 8— гиппокампальная борозда 32 ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ 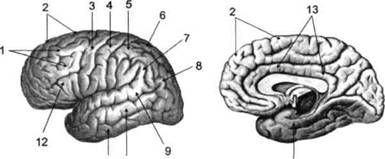 Рис. 17. Основные извилины больших полушарий головного мозга: 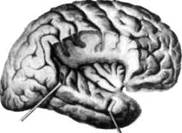 Рис. 18. Островок / — средняя лобная; 2 — верхняя лобная; 3 — передняя центральная; 4— задняя центральная; 5 — нижняя теменная долька; 6 — верхняя теменная долька; 7— надкраевая; 8 — угловая; 9 — верхняя височная; 10 — средняя височная; // — нижняя височная; 12 — нижняя лобная; 13 — поясная; 14— морского коня (гиппокампальная или парагиппокампапьная) Долей каждого полушария пять: лобная, теменная, височная, затылочная и долька, скрытая на дне сильвиевой борозды, — островок (рис. 18). Он прикрыт частями лобной, теменной и височной долей, совокупно составляющими покрышку (operculum). Оба полушария объединены между собой рядом спаек, наиболее крупная из которых — мозолистое тело — расположена выше таламуса. Общая композиция анатомических компонентов больших полушарий и нижележащих структур видна на рисунках горизонтального и вертикального срезов мозга (рис. 19 и 20). Между передней частью мозолистого тела и сводом (двухканальной системы волокон, которая проходит над таламусом и «рогообразно» соединяет гиппокамп — «морского коня» — валикоподобный выступ на медиальной стенке нижнего рога бокового желудочка, и гипоталамус) натянута тонкая вертикальная пластинка мозговой ткани — прозрачная перегородка. Совокупность отделов мозга, включающая преимущественно внутреннюю поверхность полушарий и их глубокие структуры, объединенные по пространственному и функциональному признакам, получила название лимби-ческой системы. Общепринятого представления о ее составляющих элементах не существует, границы достаточно расплывчаты и изменчивы, а четкая функциональная определенность отсутствует. Обычно предполагается большинство промежуточных образований, которые являются посредником между гипоталамусом и корой и представляют собой остаток древних нервных структур: гиппокамп, перегородка, передние ядра таламуса, свод, а также 33 ОСНОВЫ НЕЙРОПСИХОЛОГИИ 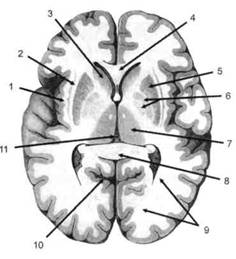 Рис. 19. Горизонтальный срез больших полушарий головного мозга: / — самая наружная капсула; 2 — наружная капсула; 3 — головка хвостатого ядра; 4 — передняя часть мозолистого тела; 5 — скорлупа; 6 — бледный шар; 7— таламус; 8 — задняя часть мозолистого тела; 9 — зрительное сияние; 10 — шпорная борозда; 11 — третий желудочек 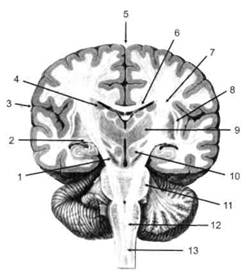 Рис. 20. Вертикальный срез головного мозга: / — черная субстанция; 2 — зрительный тракт; 3— сильвиева борозда; 4— хвост хвостатого ядра; 5— продольная щель мозга; 6— мозолистое тело; 7— внутренняя капсула; 8 — наружная капсула; 9— таламус; 10— красное ядро; //— варолиев мост; 12— пирамиды; 13 — перекрестье пирамид 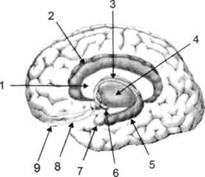 Рис. 21. Основные фрагменты лимбической системы: 1 — перегородка; 2— поясная извилина; 3 — свод; 4 —таламус; 5 — гиппокамп; 6 — мамиллярные тела; 7— миндалевидное тело (миндалина); 8— зрительный тракт; 9— зрительные луковицы мамиллярные тела (составную часть гипоталамуса) и др. В толще височных долей располагается традиционно включаемое в состав лимбической системы миндалевидное тело (миндалевидный комплекс), которое многие авторы описывают как утолщение височной коры (рис. 21). Иногда миндалевидное тело структурно причисляют к группе базаль-ных ядер. На ранних этапах эволюции многие из указанных анатомических элементов имели отношение к обонятельной функции, но позднее у высших млекопитающих почти ее утратили. Кроме вышеназванных структур в лимбическую систему в настоящее время также включают гипоталамус и ретикулярную формацию среднего мозга. ГЛАВА 2. AHATOMO-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ Особенностью лимбической системы является то, что между ее составляющими имеются простые двухсторонние связи и сложные пути, образующие множество замкнутых кругов. Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым сохранения в ней единого состояния, а также навязывания его другим системам мозга. Круги разного функционального назначения связывают лимби-ческую систему со многими структурами большого мозга, что позволяет ей реализовать функции, специфика которых определяется включенной дополнительной функциональной системой или структурой. Обилие связей лимбической системы со многими компонентами ЦНС не позволяет выделить отдельную функцию мозга, в которой она не принимала бы участие. Наиболее полифункциональными образованиями ее являются гиппокамп и миндалина. Специфических афферентных входов лимбическая система не имеет и поддерживает свою активность за счет многосторонних связей с ретикулярной формацией. Примерной условной границей лимбической системы сверху можно считать поясную извилину, дугообразно сверху примыкающую к мозолистому телу, а нижней — парагиппокампальную извилину на медиальной поверхности височных долей (рис. 22). Подавляющее большинство структур лимбической системы принимает участие в функциональной организации эмоций, что предполагает их влияние на соответствующие вегетативные изменения, регулируемые гипоталамусом, а также привлечение потенциала генетической и оперативной памяти. Кора головного мозга является наиболее дифференцированным отделом центральной нервной системы (ЦНС) и по своему морфологическому строению делится на шесть слоев, отличающихся по строению и расположению нервных элементов (рис. 23). Основными типами нейронов коры являются пирамидные, звездчатые (астроциты) и веретенообразные нейроны. В афферентной функции коры и в процессах переключения возбуждения на соседние нейроны основная роль принадлежит звездчатым нейронам. Эти клетки имеют короткие ветвящиеся аксоны, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендриты (астроцит можно увидеть в левом верхнем углу рис. 2 и в верхнем слое на рис. 23). Звездчатые нейроны участвуют в процессах восприятия, раздражении и объединении деятельности различных пирамидных нейронов. Пирамидные нейроны осуществляют эфферентную функцию коры и внут-рикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются эфферентные пути к подкорковым образованиям, и мелкие пирамиды, образующие 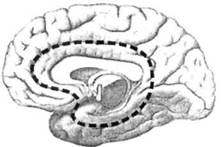 Рис. 22. Условные границы лимбической системы на медиальной поверхности полушарий ОСНОВЫ НЕЙРОПСИХОЛОГИИ 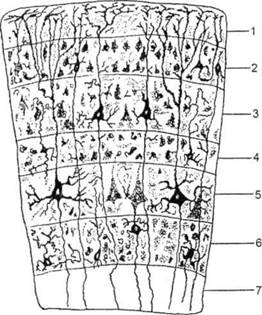 Рис. 23. Схема строения коры головного мозга: / — молекулярный (зональный) слой; 2— наружный зернистый слой; 3 — слой малых и средних пирамид; 4— внутренний зернистый слой; 5— ганглиозный слой; б — слой полиморфных клеток; 7 — белое веществою. (По М. Г. Привесу.) В пятом слое хорошо видны пирамидные клетки связи с другими отделами коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца [Владимир Алексеевич Бец] — находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид — их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит, через который в клетку поступают различные афферентные влияния от других нейронов, а вертикально вниз отходит эфферентный отросток — аксон (рис. 23, пятый слой). Многочисленность контактов (например, только на дендритах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейронов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма. Веретенообразные клетки, характерные для наружного и внутреннего зернистых слоев, в основном выполняют ассоциативную функцию — сравнительными исследованиями 28 видов приматов и человека показано, что чем ближе к Homo sapiens находится человекообразная обезьяна, тем выше в ее мозге концентрация крупных веретенообразных нейронов. ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ Прямые физиологические исследования коры доказали, что ее основной структурно-организующей единицей является так называемая кортикальная колонка, представляющая собой вертикальный нейронный модуль, все клетки которого (около 110) имеют общее рецепторное поле или однородную функциональную ориентированность (Маунткасл В. [Mountcastle Vernon]). Это сложная обрабатывающая и распределяющая единица, которая связывает ряд входов с несколькими выходами. Колонки группируются в более сложные образования — макроколонки, сохраняющие определенный топологический порядок и образующие строго связанные распределенные системы. Для коры больших полушарий характерно обилие межнейронных связей, которые по мере развития мозга человека после его рождения особенно интенсивно увеличиваются до 18 лет. Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или ее ци-тоархитектонику. Благодаря исследованиям К. Бродмана (1909), С. и О. Фогт [Cecile and Oscar Vogt, 1919-1920] и работам сотрудников Московского института мозга было выявлено более 50 различных участков коры — корковых цито-архитектонических полей, в которых нервные элементы имеют свою морфологическую и функциональную специфику (а с учетом волоконного строения коры — 150 участков). Из этих полей, обозначаемых номерами (и дополнительно буквами), составлена карта мозговой коры человека (рис. 24, 25, 26). Выделяют 52 основных поля в 11 областях коры больших полушарий головного мозга (13, 14, 15, 16, 27, 49, 50, 51 поля — только у обезьян).
Имеющаяся у современного человека кора несет на себе отпечаток филогенетического развития, благодаря чему она подразделяется на 4 «территории», отличающиеся дифференцированностью нейронных структур и их слоев: палеокортекс — древнюю кору, имеющую отношение к обонятельным функциям — обонятельную луковицу, обонятельный тракт, продырявленное вещество, обонятельные бугорки, прозрачную перегородку, латераль- ОСНОВЫ НЕЙРОПСИХОЛОГИИ 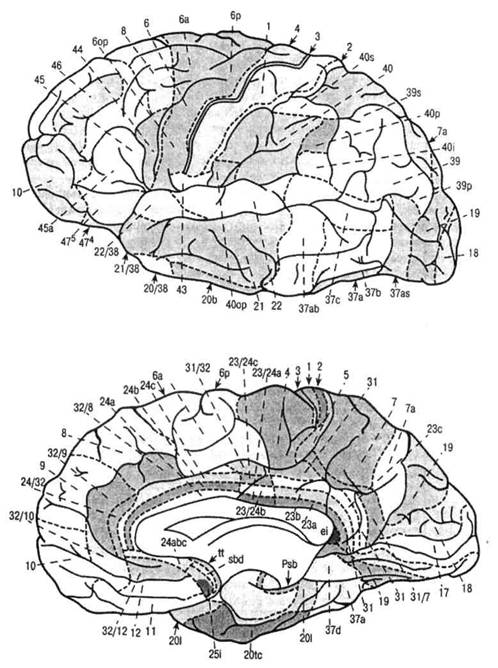 Рис. 24. Карта цитоархитектонических полей Бродмана — латерально-конвекситальные и медиальные поверхности коры (по Р. Д. Синельникову, Я. Р. Синельникову) ГЛАВА 2. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ 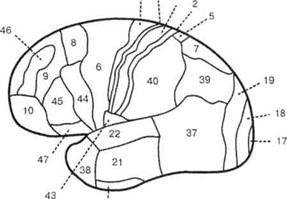 20 Рис. 25. Упрощенный вариант карты цитоархитектонических полей латерально-конвекситальной коры ную обонятельную извилину и прилежащую часть медиальной поверхности височной доли (препириформную кору), см. рис. из раздела 4.6.; архекор-текс — старую кору, сформировавшуюся позднее палеокортекса, включающую участки, расположенные на медиальной поверхности полушарий вокруг мозолистого тела и нижнего рога бокового желудочка (поясную извилину, гиппокамп, зубчатую извилину, миндалевидное тело); мезокортекс — про- 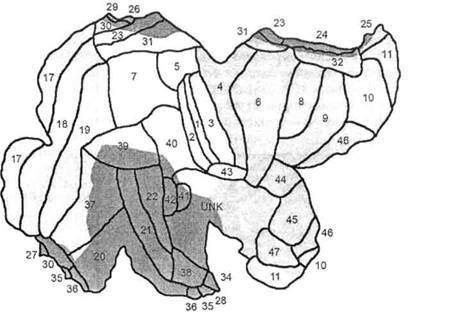 Рис. 26. Плоскостная развертка цитоархитектонических полей (по D. Van Essen, H. Drury) ОСНОВЫ НЕЙРОПСИХОЛОГИИ межуточную кору, включающую наружно-нижнюю поверхность коры островка головного мозга (иногда к мезокортексу причисляют поясную и пара-гиппокамальную извилины); неокортекс — новую кору, появляющуюся лишь у млекопитающих и составляющую около 85% коры больших полушарий, в основном лежащей на их конвекситально-латеральных (верхне-боковых) поверхностях. Палеокортекс и архиокортекс являются составными частями лимбической системы. Кора головного мозга, подкорковые структуры, а также периферические компоненты организма связаны несколькими типами проводящих путей (волокнами нейронов). Существуют различные способы классификации этих путей, наиболее общий из которых предусматривает пять их вариантов. Основной смысл подобной схемы связан с тезисом, в соответствии с которым различные типы волокон являются представителями различных систем мозга, обеспечивающими разнообразный психофизиологический эффект их работы. Ассоциативные волокна проходят внутри только одного полушария и связывают соседние извилины в виде коротких дугообразных пучков либо кору различных долей, что требует более длинных волокон. Назначение ассоциативных связей — обеспечение целостной работы одного полушария как анализатора и синтезатора разномодальных (имеющих отношение к разным сенсорным системам) возбуждений (рис. 27). Проекционные волокна связывают периферические рецепторы с корой головного мозга. Они имеют самый разный ход внутри ЦНС, зависимый от той системы, в состав которой входят. Подавляющая часть проекционных волокон перекрещивается на разных уровнях головного или спинного мозга и почти для всех из них (за исключением обонятельных) промежуточной инстанцией на пути к коре становится таламус. По характеру своей роли это афферентные пути, задача которых — трансляция мономодального импульса к соответствующим корковым представительствам того или иного анализатора. Интегративно-пусковые волокна начинаются от двигательных зон мозга, являются нисходящими эфферентными и по аналогии с проекционными также имеют перекрестья на различных уровнях стволового участка или спинного мозга. Задача этих волокон — трансляция возбуждений разных модальностей в це- 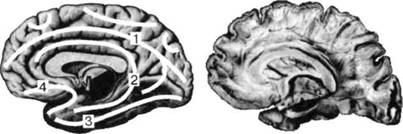 Рис. 27. Схема основных пучков ассоциативных волокон: / — верхний продольный; 2— поясной; 3— нижний продольный; 4— крючковидный. Справа на препарате хорошо виден ход некоторых волокон 40 ГЛАВА 2. AHATOMO-ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ ленаправленно организованную или неосознаваемую двигательную активность. Окончательной зоной приложения интегративно-пусковых волокон является мышечный аппарат человека. Сточки зрения их топологической организации они также могут рассматриваться и как проекционные, поскольку реализуют принцип строгого соответствия (фактически — связи) между центральными корковыми нейронными группами и периферическими мышечными волокнами. Комиссуральные волокна обеспечивают целостную совместную работу двух полушарий. Они представлены одним крупным анатомическим образованием — мозолистым телом и несколькими более мелкими структурами, важнейшими из которых являются четверохолмие, зрительная хиазма и межуточная масса таламуса (место соединения парныхталамических образований) (рис. 9, 19, 20, 28). Функционально мозолистое тело состоит из трех отделов: переднего, среднего и заднего. Его поперечные волокна, вступив в полушарие, радиально расходятся и образуют лучистость мозолистого тела соответственно долям мозга. Волокна мозолистого тела идут в разных направлениях, но главным образом — в поперечном, связывая симметричные места полушарий и осуществляя между ними обмен нервной импульсацией. В нем также проходят волокна, соединяющие разноимённые извилины противоположных полушарий. Передний отдел мозолистого тела преимущественно обслуживает процессы взаимодействия в двигательной сфере, средний — в слуховой, слухоречевой и кинестетической, а задний — в тактильной и зрительной. Предположительно большая часть волокон мозолистого тела участвует в межполу-шарных ассоциативных процессах, регуляция которых может сводиться как к взаимной активации объединяемых участков мозга, так и к торможению деятельности контралатеральных (с противоположной стороны) зон. Лимбико-ретикулярные волокна связывают энергорегулирующие зоны продолговатого мозга с корой. Основная задача этих путей — поддержание циклов общего активного или пассивного фона мозга, выражающихся для человека в феноменах бодрствования, ясного сознания или сна, а также в более дифференцированных влияниях на кору больших полушарий. В качестве систем, способных управлять энергетическим потенциалом ЦНС и всего организма, называются две — ретикулярная формация и лимбическая система. Область распространения ретикулярной формации точно не установлена. Морфологические и физиологические данные говорят о том, что она занимает центральное положение в продолговатом мозгу, мосту, среднем мозгу, в гипотала-мической области и даже в некоторых участках зрительных бугров. Восходящие волокна ретикулярной формации изнутри разбрасываются по всей коре, но наиболее мощные связи продолговатого мозга с корой отслеживаются в отношении лобных долей (рис. 29). Определенная 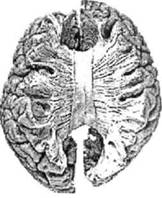 Рис. 28. Мозолистое тело с лучистой короной (препарат) ОСНОВЫ НЕЙРОПСИХОЛОГИИ 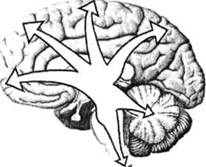 Рис. 29. Схема распространения влияния ретикулярной формации часть нисходящих ретикулярных волокон обслуживает и работу спинного мозга. Через эти пути ретикулярная формация может избирательно активизировать или тормозить работу зон мозга, в которые они направляются. Непосредственным источником моти-вационного возбуждения являются гипо-таламические структуры, в которые поступает информация о текущем состоянии различных систем организма и о нарушениях в их деятельности. Первая инстанция, куда адресуется возбуждение любого мотивационного центра гипоталамуса, — это лимбическая система мозга, через волокна которой активируются висцеральные, эмоциональные и поведенческие реакции, направленные на восстановление того нарушенного равновесия, которое было отражено в состоянии гипоталамуса. Ретикулярная формация и лимбическая система, будучи модулирующими (меняющими параметры), тесно связаны с высшими отделами коры больших полушарий и образуют особый функциональный блок, имеющий несколько уровней реагирования: физиологический, психологический и поведенческий. В соответствии с этой логикой, функциональное состояние человека можно рассматривать как результат активности этой объединенной системы и ее волокон. 2.3. СТРУКТУРНЫЙ ФИЛО-И ОНТОГЕНЕЗ МОЗГА ЧЕЛОВЕКА Мозг человека как специальный орган, осуществляющий высшую форму обработки информации, представляет собой лишь часть нервного аппарата — системы, специализирующейся на согласовании внутренних потребностей организма с возможностями их реализации во внешней, в том числе социальной, среде. Как и всякая система, она имеет определенную пространственную и функциональную конструкцию, сформировавшуюся в ходе эволюционного процесса. Поэтому диапазон основных параметров функционирования нервной системы в целом отражает вероятностную структуру качества и интенсивности раздражителей, с которыми формирующийся организм сталкивался на протяжении фило- и онтогенеза. В широком плане нервная система с входящим в нее мозгом — это иерархически и функционально упорядоченное материальное пространство, являющееся неотъемлемым элементом еще более общей системы — организма. 42 |