Цикл трикарбоновых кислот
 Скачать 3.84 Mb. Скачать 3.84 Mb.
|

|
Глюкозо-1-фосфат + УТФ < = > УДФ-глюкоза +Пирофосфат. 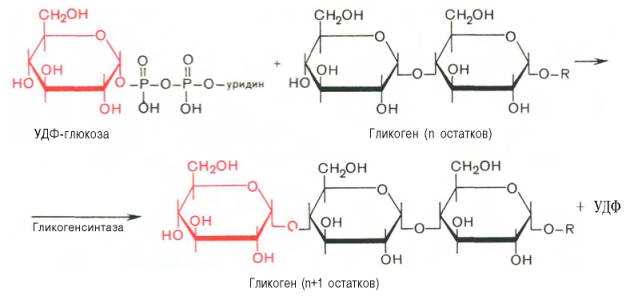 Эта реакция катализируется ферментом гликогенсинтазой. Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ+. 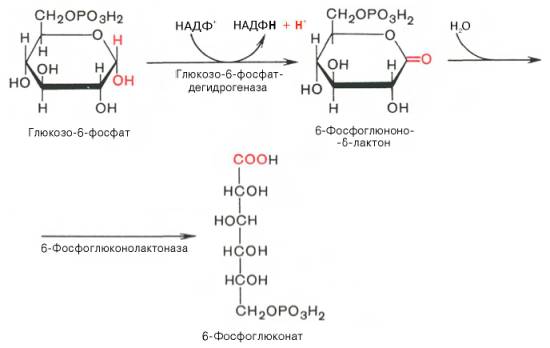 Во второй – окислительной – реакции, катализируемой 6-фосфоглюко-натдегидрогеназой (декарбоксилирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН: 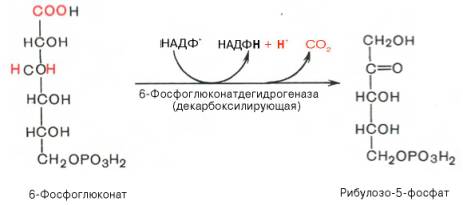 Анаэробный гликолиз у животных и человека может происходить во многих типах клеток, но его значении для разных органов различно. Особенное большое значение он имеет при кратковременной интенсивной работе, так бег в течение 30 с обеспеч анаэробным гликолизом. Эритроциты вообще не имеют митохондрий и их потребность в атф удовлетвор за счет анаэробного гликолиза. Билет 7 1. Ферменты цикла Кребса и тканевого дыхания локализованы во фракции митохондрий. 2. 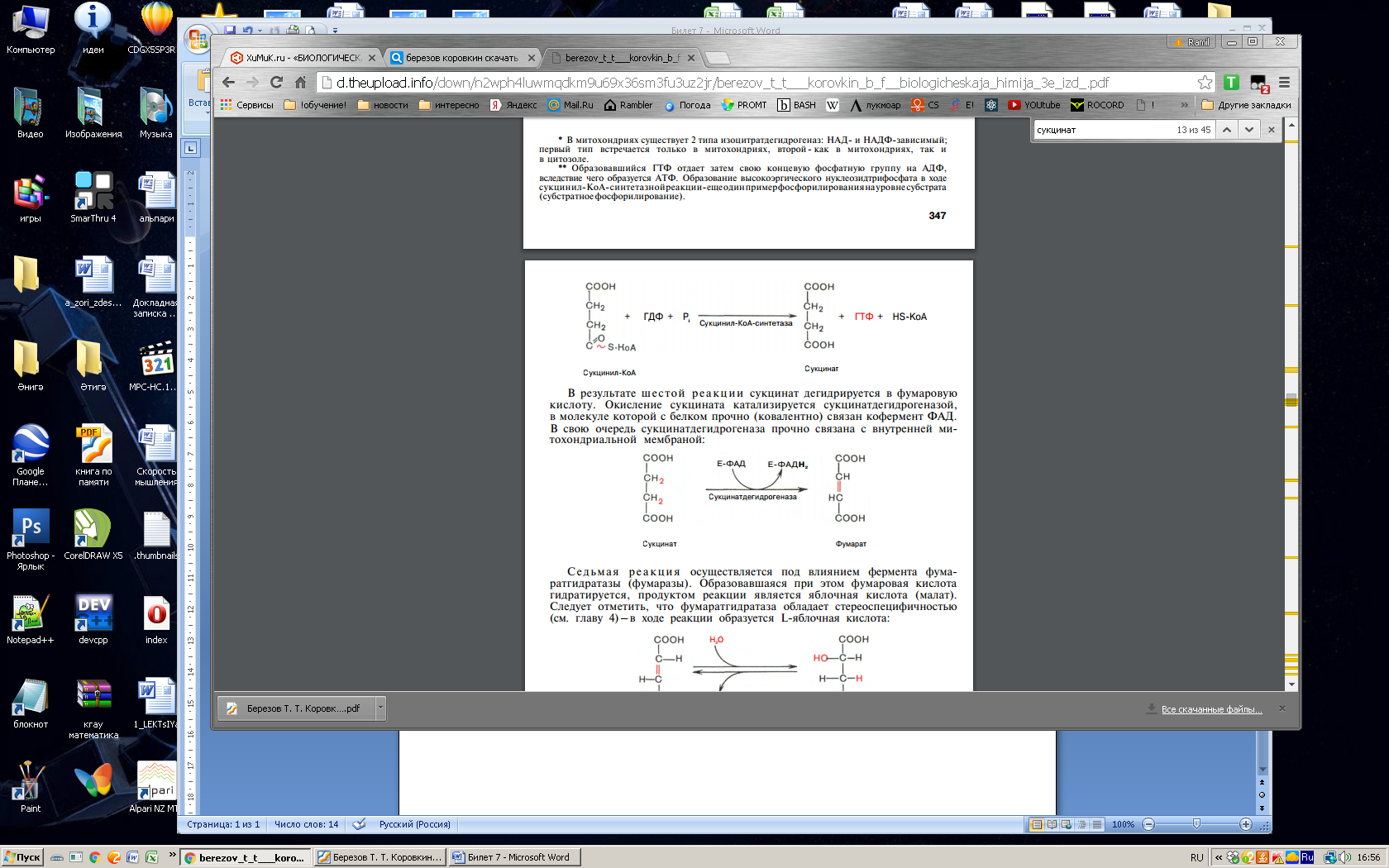 Фермент – сукцини-КоА-синтетаза 3. Мобилизация (распад) гликогена. ?Фермент? 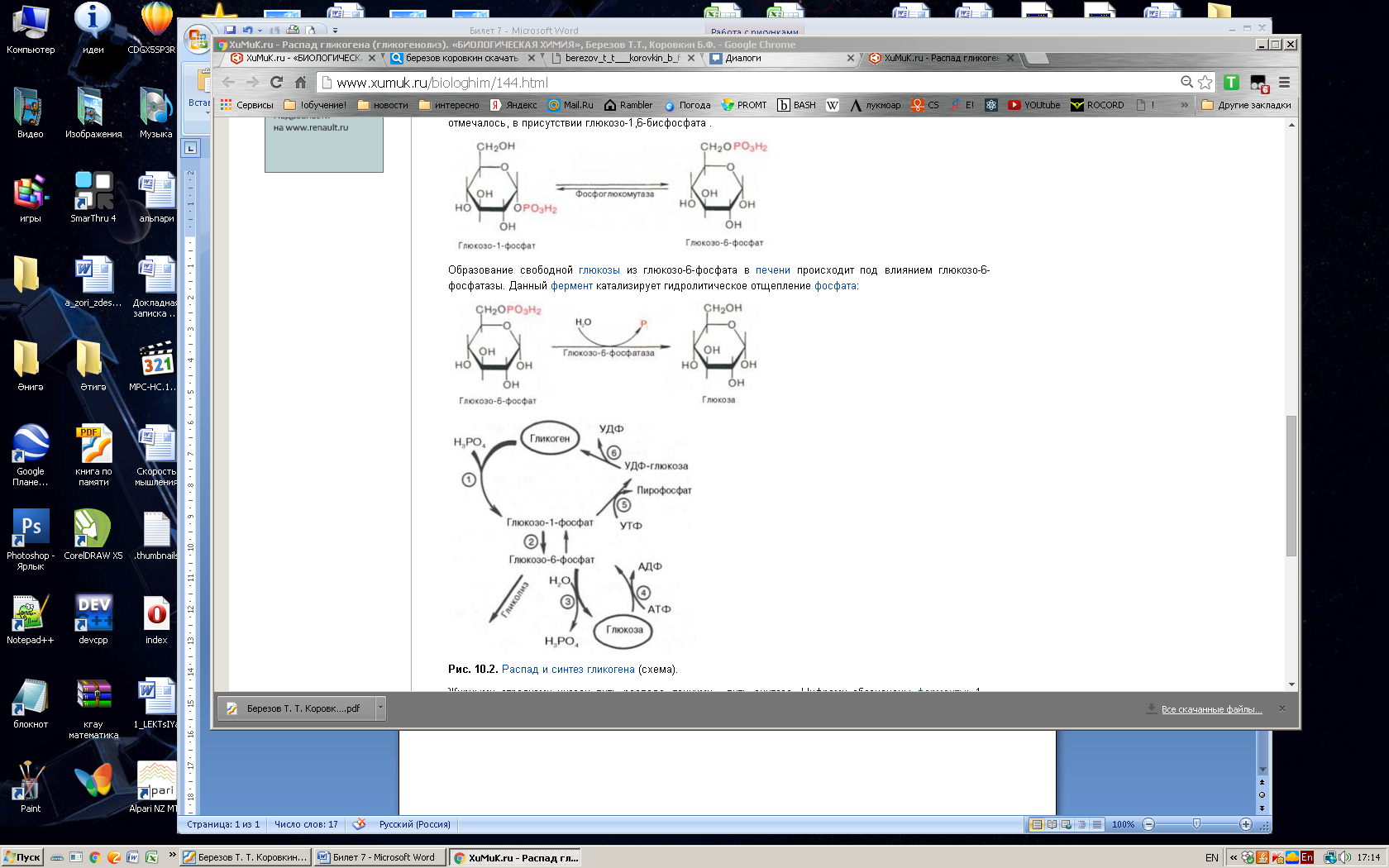 Гормональная регуляция 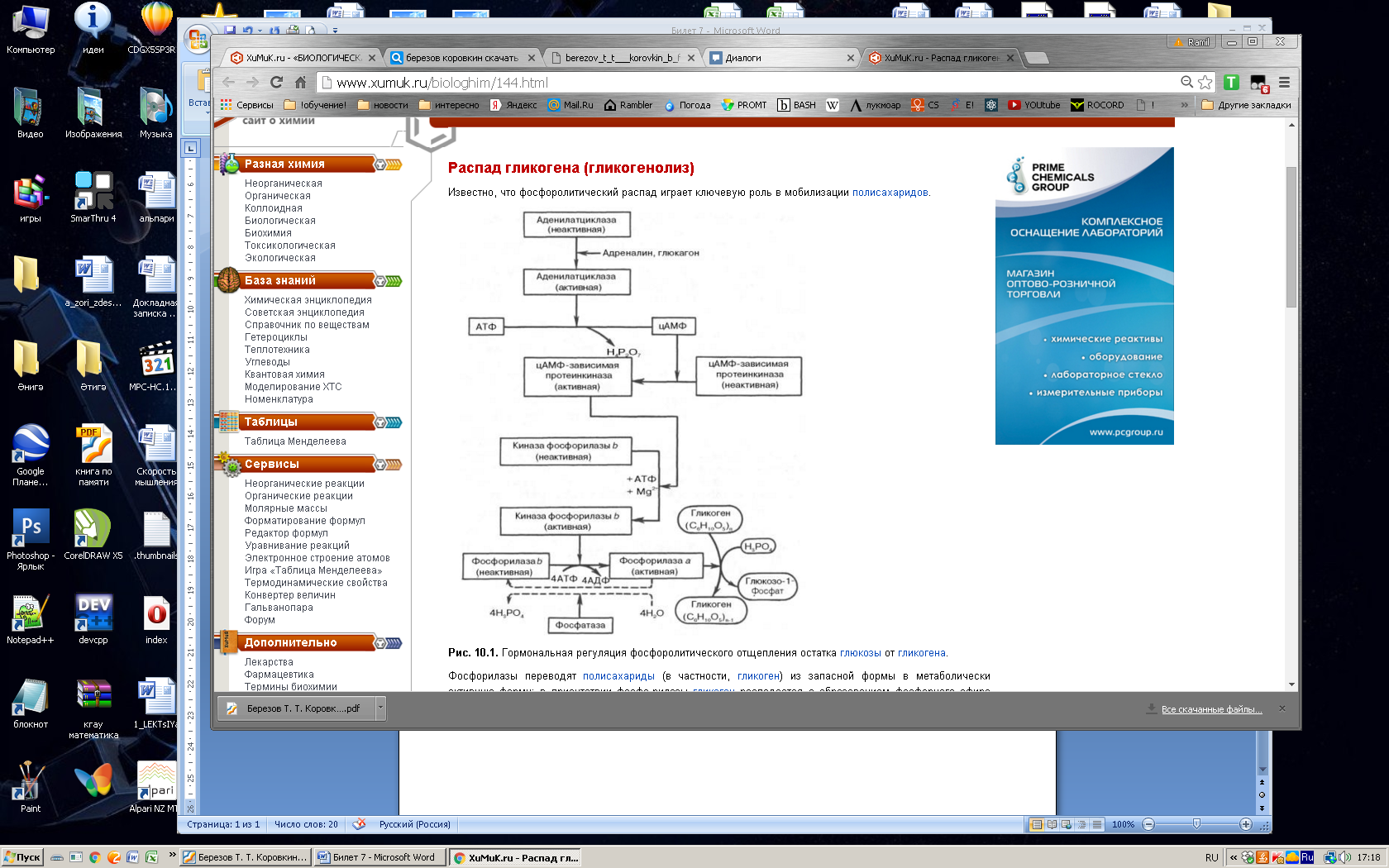 4. Аэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных в присутствии кислорода, включает те же стадии, что и анаэробный гликолиз, за исключением последней стадии: восстановления пирувата до лактата, которая протекает в анаэробных условиях Специфические характеристики аэробного гликолиза (отличие от анаэробного гликолиза): Во-первых, окисление восстановленной формы НАДН∙Н+, являющееся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. Во-вторых, образование АТФ при аэробном гликолизе может идти двумя путями: - путём субстратного фосфорилирования, когда для фосфорилирования АДФ используется энергия макроэргической связи субстрата; - путем окислительного фосфорилирования АДФ, сопряженного с дыхательной цепью. 5. Глюкозо-лактатный цикл (цикл Кори) – это циклический процесс, объединяющий реакции глюконеогенеза и реакции анаэробного гликолиза. Глюконеогенез происходит в печени, субстратом для синтеза глюкозы является лактат, поступающий в основном из эритроцитов или мышечной ткани. В эритроцитах молочная кислота образуется непрерывно, так как для них анаэробный гликолиз является единственным способом образования энергии. В скелетных мышцах высокое накопление молочной кислоты (лактата) является следствием гликолиза при очень интенсивной, субмаксимальной мощности, работе, при этом внутриклеточный рН снижается до 6,3-6,5. Но даже при работе низкой и средней интенсивности в скелетной мышце всегда образуется некоторое количество лактата. Убрать молочную кислоту можно только одним способом – превратить ее в пировиноградную кислоту. Однако сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в пируват из-за особенностей изофермента лактатдегидрогеназы-5. Зато клеточная мембрана высоко проницаема для лактата и он движется по градиенту концентрации наружу. Поэтому во время и после нагрузки (при восстановлении) лактат легко удаляется из мышцы. Это происходит довольно быстро, всего через 0,5-1,5 часа в мышце лактата уже нет. Малая часть молочной кислоты выводится с мочой. Большая часть лактата крови захватывается гепатоцитами, окисляется в пировиноградную кислоту и вступает на путь глюконеогенеза. Глюкоза, образованная в печени используется самим гепатоцитом или возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена. Также она может распределиться по другим органам. Билет 8 Гликогенозы.-ряд наследственных болезней связанные с нарушением обмена Гликогена. Они возникаютв связи с дефицитом или полным отсутствием ферментов, катализирующихпроцессы распада или синтеза гликогена, и характеризуются избыточным его накоплением в различных органах и тканях. Агликогеноз - наследственное заболевание, вызванное отсутствием фермента, ответственного за синтез гликогена, а именно уридин-дифосфат-глюкозо-гликогентрансферазы или гликогенсинтетазы. 2. Гликемический профиль – это динамическое наблюдение за уровнем сахара в крови в течение суток. Обычно производят 6 или 8 заборов крови из пальца для определения уровня глюкозы: перед каждым приемом пищи и через 90 минут после еды. Для сахарного диабета I типа уровень глюкозы считается компенсированным, если ее концентрация натощак и в течение суток не превышает 10 ммоль/л. Для данной формы заболевания допустима небольшая потеря сахара с мочой – до 30 г/сут. Сахарный диабет II типа считается скомпенсированным, если концентрация глюкозы в крови утром не превышает 6,0 ммоль/л, а в течение дня – до 8,25 ммоль/л. Глюкоза в моче определяться не должна. У здорового человека норма уровня сахара в крови составляет: -натощак:от 3,5 до 5,7 ммоль/л; -через два часа после приема пищи: от 4,5 до 7,0 ммоль/л; - ночью: от 3,9 до 5,7 ммоль/л. 3. Глюконеогенез это процесс образования глюкозы из органических соединений, в печени,а также в корковом веществе почек и в эпителии тонкого кишечника. 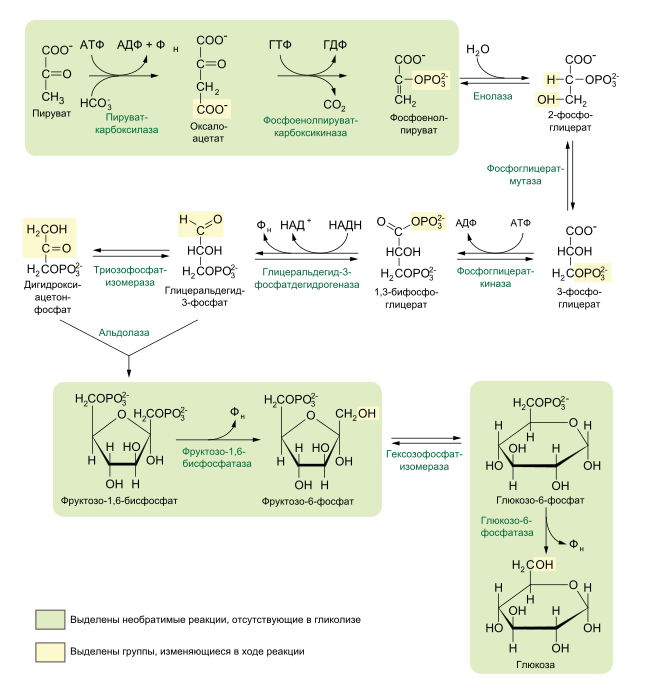 Гормональная активация глюконеогенеза осуществляется глюкокортикоидами, которые увеличивают синтез пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы,фруктозо-1,6-дифосфатазы. Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм путем фосфорилирования. Также имеется метаболическая регуляция, при которой аллостерически активируетсяпируваткарбоксилаза при помощи ацетил-SКоА, фруктозо-1,6-дифосфатаза при участии АТФ. 4. Анаэробный гликолиз. происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН  Аэробный гликолиз 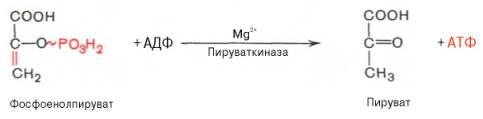 5. 12 молекул АТФ Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ. Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ. Билет 9 болезнь Гирке, возникает в результате дефекта глюкозо-6-фосфатазы. 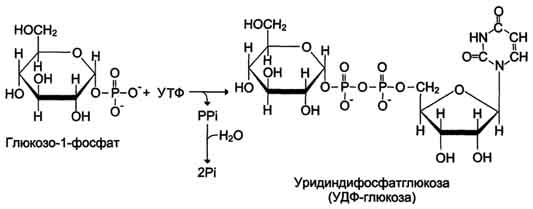 Переваривание углеводов начинается уже в ротовой полости. В слюне присутствует гидролитический фермент α-амилаза, расщепляющая в крахмале α-1,4-гликозидные связи.В ротовой полости не может происходить полное расщепление крахмала, так как действие фермента на крахмал кратковременно. Крахмал переваривается лишь частично с образованием мальтозы. Мальтоза является временным продуктом, так как она гидролизируется под влиянием фермента мальтазы на молекулы глюкозы. Кишечный сок так же содержит активную 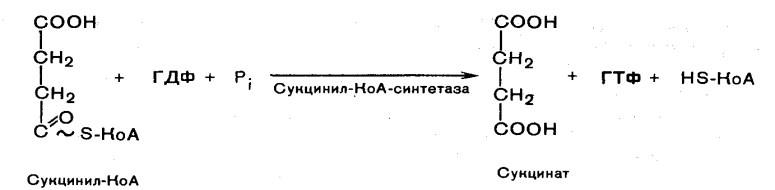 сахаразу под влиянием которой из сахарозы образуется глюкоза и сахароза. сахаразу под влиянием которой из сахарозы образуется глюкоза и сахароза.Скорость всасывания моносахаридов различна. Принято считать, что всасывание маннозы, пентозы и фруктозы осуществляется преимущественно путём диффузии, всасывание других моносахаридов за счет активного транспорта. Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. 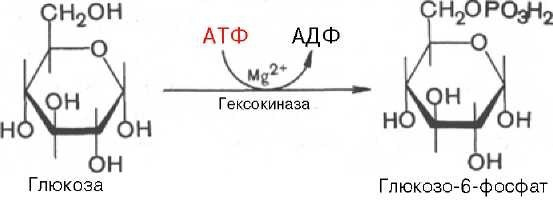 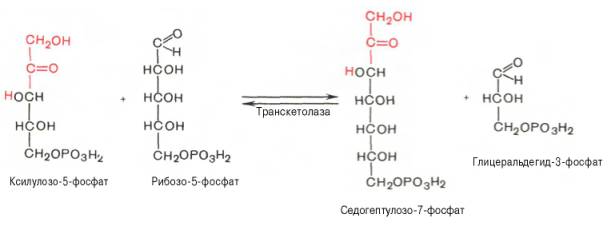 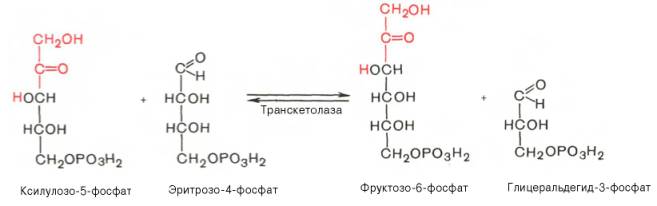 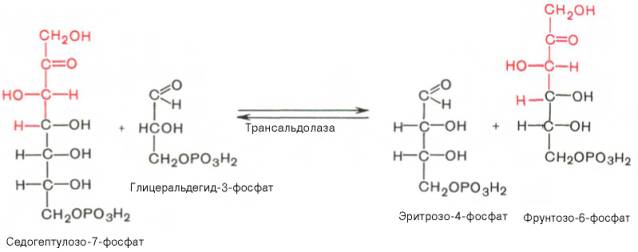 38 АТФ образуется при полном окислении одной молекулы глюкозы до СО2 и воды БИЛЕТ 10 1. В каких реакциях цикла Кребса участвует КоА? В образовании цитрат – иона участвует ацетил – КоА 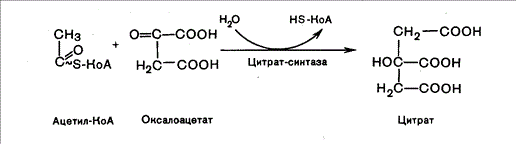 окисление α-кетоглутарата до сукцинил-СоА 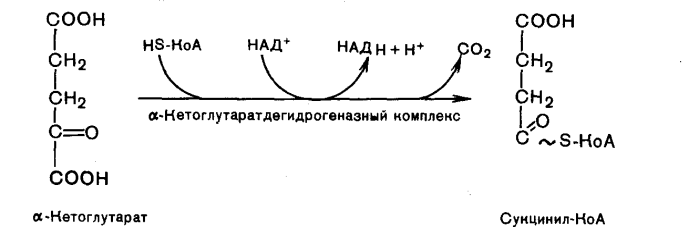 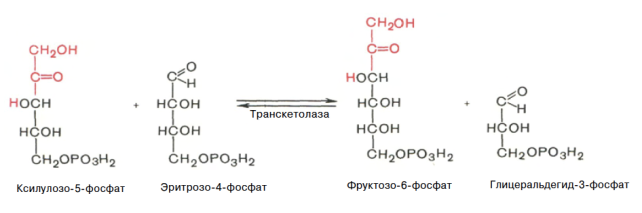 превращение сукцинил-СоА в сукцинат. 2. Напишите реакцию окислительного фосфорилирования в процессе гликолиза. Укажите фермент. 3. Напишите реакции окислительного декарбоксилирования пирувата. Укажите ферменты. В какой части клетки протекает этот процесс? Каково строение пируватдегидрогеназного комплекса? Этот процесс протекает в матриксе митохондрий. Ферменты: пируватдегидрогиназа, 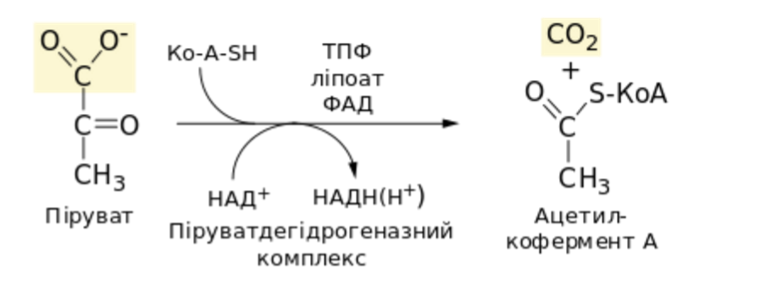 дигидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа. дигидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа.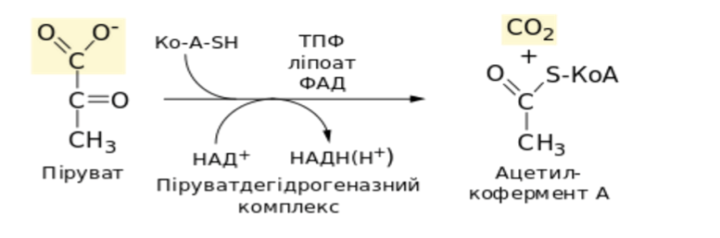 Пируватдегидрогеназный комплекссостоит из 3 ферментов: Пируватдегидрогеназный комплекссостоит из 3 ферментов:пируватдекарбоксилаза (Е1), дигидролипоилтрансацетилаза (Е2), дигидролипоилдегидрогеназа (Е3). 4. Напишите реакции неокислительной стадии гликолиза. Укажите ферменты. Сколько АТФ потребляется на этой стадии? 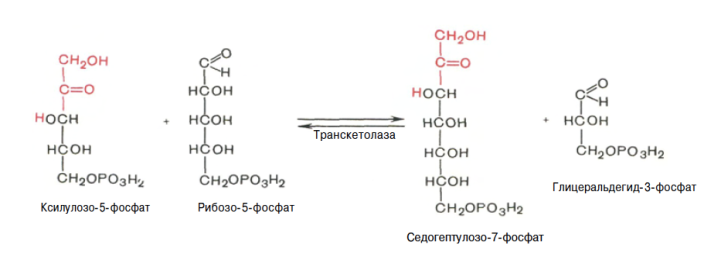 5. Сколько АТФ образуется при полном окислении одной молекулы пирувата до СО2 и воды? 15 молекул АТФ. Билет 11 Почеченый порог- наличие глюкозы в моче. В норме моча не содержит глюкозы, посколькупочки способны реабсорбировать (возвращать в кровоток) весь объём глюкозы, прошедший через почечный клубочек в просвет канальцев нефрона. В подавляющем большинстве случаев гликозурия является сипмтомом декомпенсированного сахарного диабета как результат патологического увеличения концентрации глюкозы в крови. Гликозурия ведёт к избыточной потере воды с мочой — обезвоживанию организма, развивающемуся из-за усиления осмотического компонента диуреза. У здоровых людей почеченый порог 170-180 мг/100мл. 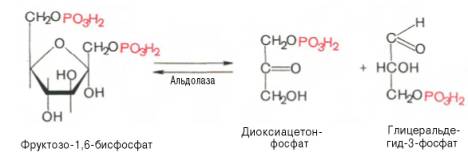 Ранеостатическое постоянство уровня глюкозы в крови обусловлено тем, что она подвергается регулированию. - регуляция гомеостаза инсулином высвобождается в ответ на увеличение глюкозы -регуляция гомеостаза глюкозы глюкагоном. На снижение содержания глюкозы в крови секретируется глюкагон -регуляция адреналином, глюкокортикостероидами, соматотропином, йодтиронином 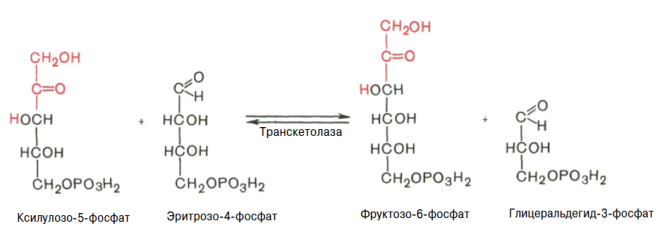 Норм конц в крови 3,3- 5,5 ммоль/л Норм конц в крови 3,3- 5,5 ммоль/лСтадия гликолитической оксидоредукции 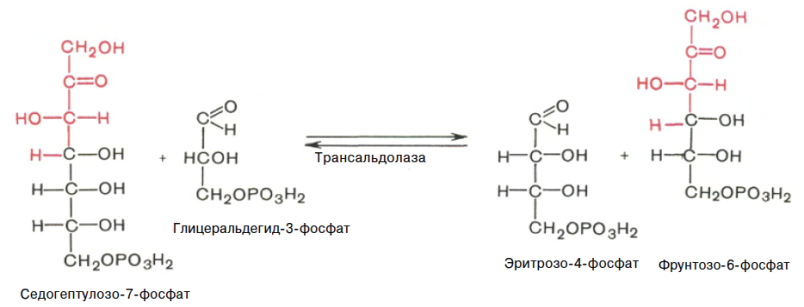 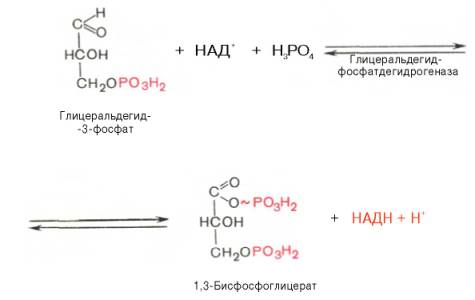  Надн=2,5 атф 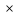 2=5 атф образовалось 2=5 атф образовалосьВ местной реакции анаэробного гликолиза в процессе образования НАДН2 глицеральдегидтрифосфат связывается с молекулой фермента за счет SH группы . образ связь богатая энергией но она не прочная и расщепляется под влиянием неорганического фосфата и образуется 1,3 дифосфоглицерат. Билет12 Органами-мишенями для глюкагона являются печень, миокард, жировая ткань. В печени гормон ускоряет мобилезацию гликогена вызывая торможения гликолиза, стимулирует гликонеогенез. В жировой ткани ускоряет мобилизацию триацетилглицеролов. В корковом веществе почек активирует глюконеогенез. 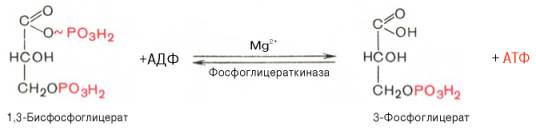 В первой реакции, катализируемой ферментом цитратсинтазой, ацетил-КоА конденсируется с оксалоацетатом. В результате образуется лимонная кислота: 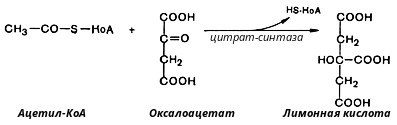 Катализирует эти обратимые реакции гидратации- дегидратации фермент аконитат-гидратаза: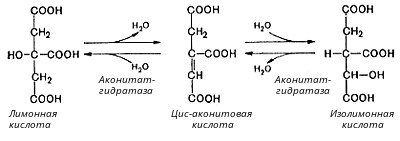 В третьей реакции, в присутствии НАД-зависимой изоцитратдегидрогеназы: 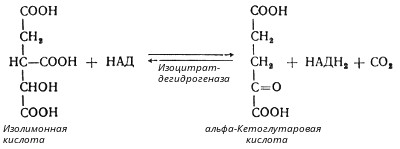 В четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты до сукцинил-КоА. 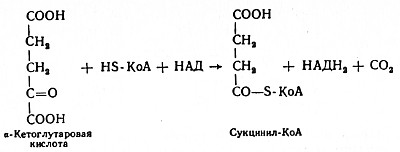 Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. 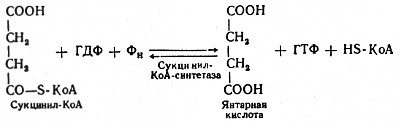 В шестой реакции Окисление сукцината катализируется сукцинатдегидрогеназой, 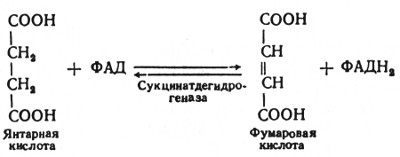 В седьмой реакции под влиянием фермента фумаратгидратазы. 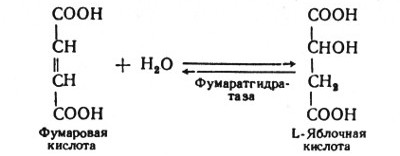 Наконец, в восьмой реакции под влиянием митохондриальной НАД-зависимой малатдегидрогеназы 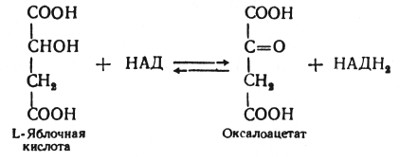 1НАДН=2,5АТФ (обр-сь 3шт) 2,5*3= 7,5+1+1,5=10АТФ (всего) ФАДН=1,5 АТФ ГТФ=1АТФ Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД. В цитозолеметаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H+как бы "превращается" в митохондриальный ФАДН2. Этот челнок активен в печени и белых скелетных мышцах и необходим для получения энергиииз глюкозы при работе клетки. Галактоземия – наследственное заболевания в основе которого лежит нарушения обмена обмена веществ на пути преобразования галактозыв лактозу. Это связано с нарушением стр-го гена ответственного за синтез фермента галактоза 1 фосфатуридил трансфераза. Билет13 Алифатическая регуляция скорости гликолиза зависит от изменения состояния АТФ/АДФ направленное измерения скорости использования глюкозы непосредственно в печень. Глюкоза в клетках печени используется но только для синтеза гликогена и жиров. Основными потребителями АТФ в гепатоцитах являются процессы трансмембранного комплекса , синтез белка , глюконеогенез.  В гликогенолизе непосредственно участвует три фермента:1) фосфорилаза гликоена, 2) плюкантрансфераза,3) амино альфа 1,6 глюкозидаза . Метаболизм гликогена в печени регулируется несколькими гормонами одни активируют ферменты сентазы другие распад.Основные ферменты метаболизма гликогена активны либо в фосфорилировании либо в дефосфорилированной форме.Присоединение фосфатов к ферменту производят протеинкиназы источником фосфора является АТФ. Фосфорилаза гликогеная активная после присоединения фосфатной группы.Синтаза после присоединения фосфата инак-ся. Адреналин и глюкагон вызывают гликогенализактивируя фосфорилазугликогена. Метоболизм этилового спирта включат реакции дегидрирования в результате которых образуется уксусная кислота. Примерно90% атетата образуется в печени.Часть в печени привращается в ацетил- КОА но большая часть из гепатоцитов попадает в кровь , а затем в мышцы где тоже превращается в ацетил-КОА. В результате быстрого дегидрирования больших количеств этанола в клетках печени уменьшается концентрация НАД+и увеличивается концентрация НАДН. Для окисления 125 г этанола требуется НАД" столько же, сколько для окисле ния 500 г глюкозы, т. е. такое количество углево дов, которое потребляется за сутки. Значитель ная часть глюкозы пищи после еды депонируется в форме гликогена и расходуется постепенно. Обмен этанола происходит за суще ственно более короткое время, особенно первая реакция — образование ацеталь- дегида, поэтому после приема алкоголя отношение [НАД+] / [НАДН] уменьшает ся. Это ведет к тому, что изменяется скорость всех реакций, зависящих от НАД" и НАДН. В частности, изменяются стационарные концентрации пирувата и лактата: Пируват + НАДН + H+ <->Лактат + НАД+. Концентрация пирувата в клетках и в крови уменьшается, а концентрация лактата увеличивается. При больших дозах и хроническом потреблении алкоголя в крови содержит ся много ацетальдегида, который повреждает мембранные структуры клеток. В частности, повреждаются митохондрии: снижается трансмембранный потенци ал и эффективность сопряжения дыхания с фосфорилированием. При участие фермента дигидролипоилдегидрогеназы осуществляется перенос атома водорода от восстановленных сульфгидрильных групп дегидроипоамида на ФАД который выполняет роль посреднеческой группы данного фермента и прочно связывается с ним. Востановленный ФАДН дигидролипоилдегидрогеназы передает Н+ на мкофермент НАД с образованием НАДН+Н+ 14 билет 1 Какими двумя соединениями ингибируется и чем активируется фосфофруктокиназа? Фосфофруктокиназа ингибируется цитратом и высокими концентрациями АТФ и активируется АДФ и АМФ и ионами кальция. Жирные кислоты и из производные яв-ся ингибиторами. 2. Какой из транспортеров глюкозы (ГЛЮТ-1, ГЛЮТ-2, ГЛЮТ-3, ГЛЮТ-4, ГЛЮТ-5) является инсулинзависимым. В какой части клетки он находится при отсутствии гормонального сигнала? ГЛЮТ-4 - инсулинзависимый, в мышцах (скелетной, сердечной), жировой ткани; Содержится в отсутствие инсулина почти полностью в цитоплазме. В отсутствие гормонального сигнала ГЛЮТ-4 находятся в цитозольных везикулах. Напишите реакции мобилизации гликогена. Укажите ферменты. Какова судьба глюкозо-1-фосфата и глюкозо-6-фосфата в печени и мышцах? Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Для осуществления данной реакции необходима фосфо-рилированная форма фосфоглюкомутазы, т.е. ее активная форма, которая образуется, как отмечалось, в присутствии глюкозо-1,6-бисфосфата .  Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата: 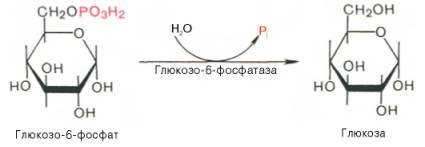 фосфорилированная глюкоза в противоположность неэте-рифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глюкозо-6-фосфатазу, который и обеспечивает возможность быстрого выхода глюкозы из этого органа. В мышечной тканиглюкозо-6-фосфатаза практически отсутствует. 4 Укажите необратимые реакции гликолиза и напишите реакции обходные путей глюконеогенеза. Укажите ферменты. Где протекают эти реакции? Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом. реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ: 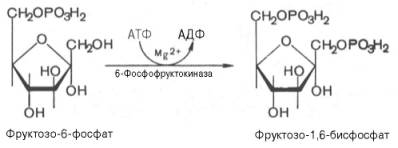 Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скоростьгликолиза в целом. Обходные пути 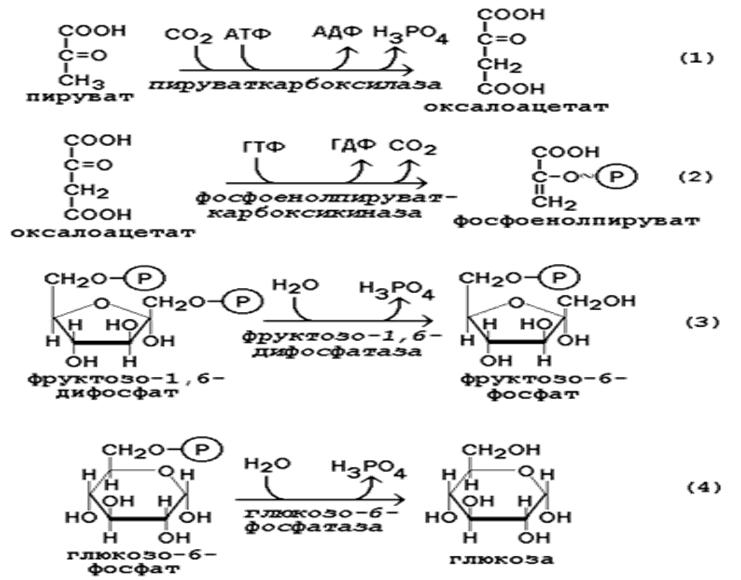 5. Какими связями соединены остатки глюкозы в молекулах амилопектина и гликогена? В линейных цепях амилопектина остатки глюкозы соединены α-1,4-связями, а в точках ветвления амилопектина – межцепочечными α-1–>6-связями. Молекула гликогена построена из ветвящихся полиглюкозидных цепей, в которых остатки глюкозы соединены α-1,4-гликозидными связями. В точках ветвления имеются α-1,6-гликозидные связи. По строению гликоген близок к амилопектину. 15 билет Инсулин. 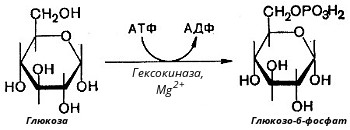 Глюкокназа,Гексокиназа Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям. Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы (АСТ) реакции трансаминированияаспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназыи за счет "гликолитического" НАДН восстанавливается дояблочной кислоты (малата). Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль. Таким образом, атомы водорода от цитозольного НАДН перемещаются в составмитохондриального НАДН. 4. Метаболизм фруктозы в клетке начинается с реакции фосфорилирования: |