ихтиопаталогия. Дана характеристика рыб как переносчиков возбудителей болезней человека и животных. Для студентов вузов по специальности Водные биоресурсы и аквакультура
 Скачать 10.48 Mb. Скачать 10.48 Mb.
|

|
Бруклинеллез Возбудитель — Brooklynella hostilis (сем. Hartmanellidae) — по строению и по характеру действия на хозяина является морским аналогом хилодонелл. Инфузория имеет фасоле- или почкообразную форму. На вентральной стороне клетки расположены ряды ресничных полос. Размеры инфузорий колеблются от 56x31 до 86x50 мкм. Паразит локализуется на жабрах рыб, вызывая многочисленные кровоизлияния, клеточную пролиферацию и другие, иногда значительные воспалительные реакции. У рыбы нарушается дыхание. В естественных водоемах у морских рыб бруклинея встречается в незначительных количествах, не вызывая каких-либо патологий у хозяина. Заболевание бруклинеллезом известно для морских рыб, содержащихся в аквариумах. Эта инфузория может стать причиной эпизоотии для рыб, выращиваемых в морских садках и бассейнах. Триходины морских рыб У морских рыб инфузории сем. Trichodinidae представлены тремя видами: Trichodina, Trichodinella и Tripartiella. Для морских рыб их описано свыше 60 видов. В отличие от пресноводных рыб у морских триходины преимущественно локализуются на жабрах. Они раздражают жаберную ткань, скользя по эпителию или прикрепляясь к нему. При нормальных условиях и слабой зараженности они не приносят ощутимого вреда хозяевам, питаясь отмершими клетками эпидермиса и элементами микрофлоры. Триходины часто присутствуют у рыб, обитающих в шельфовой зоне морей и океанов. Они известны у костных и хрящевых рыб, однако зараженность ими невелика. Считается, что процент заражения морских рыб триходинами в холодных водах гораздо выше, чем в умеренных зонах. При пересадке морских рыб в аквариумы изменения условий среды и ослабление организма могут привести к интенсивному размножению инфузорий и развитию болезни у рыб. Потенциально опасны триходины и для хозяйств марикуль-туры. При массовом заражении рыб (особенно молоди) инфузориями может произойти полное разрушение жаберного эпителия, что приводит к гибели рыб. У морских рыб широко распространена Trichodina rectuncinata, отмеченная у 15 видов рыб в различных морских бассейнах. Большинство триходин морских рыб обладают широкой специфичностью и распространены в разных географических зонах Мирового океана. Т. inversa обнаружена в Черном и Японском морях. Т. do-merguei f. partidisci встречена на рыбах Черного моря, в северо-западной и юго-западной частях Атлантического океана. Т. cotti-darum, Т. borealis, T. elegini поражают рыб в Белом и Баренцевом Морях, а также в Тихом океане. Т. borealis паразитирует также у Камбал в Черном море. Интересно, что Т. cottidarum паразитирует в яйцеводах и иногда в мочевом пузыре ската обыкновенного в Баренцевом море. Она же поражает жабры беломорского бычка (на 68 %) при высокой интенсивности инвазии. У зараженных рыб жабры становятся бледно-серыми, что свидетельствует о сильной атрофии жаберной ткани. Из инфузорий других систематических групп у морских рыб иногда встречаются сидячие инфузории родов Ambiphrya, Scyphidia, Clausophrya. Эти инфузории обнаруживают очень редко в мелководных, прибрежных зонах морей. Так, Ambiphrya miri ин-вазирует морскую иглу Nerophis ophidion в Балтийском море. Scyphidia aretica заражает Ziparis gibbus в Баренцевом море и встречается у бычков Тихоокеанского и Атлантического побережий Северной Америки. Clausophrya oblida отмечена на поверхности тела бычка-цуцика в Черном море. 8.1.6. СОСУЩИЕ ИНФУЗОРИИ РОДА CAPRINIANA Возбудитель. Сосущие инфузории из отр. Suctorida (семейства Trichophryidae) обитают на жабрах рыб. Тело трихофрий мешковидной асимметричной формы. Сосущие щупальца располагаются на переднем и заднем концах тела, прикрепляются особой секрецией. Размножаются путем множественного наружного и внутреннего почкования. Образующиеся томиты (дочерние особи) малоподвижные, стремятся осесть на жабрах того же хозяина, образуя скопления сукторий на жабрах. Все ранее описанные виды, относимые к роду Tri-chophrya, объединены в один вид из рода Capriniana — С. piscium (рис. 61). Эпизоотология. С. piscium встречаются на жабрах разных видов рыб, как в Евразии, так и в Северной Америке, у американского сомика, черного американского окуня, молоди лососей, сиговых, окуня, колюшки, у белого и черного амуров, амурской щуки, красноперки, толстолобиков и др. Заболевания отмечены в Китае у молоди толстолобиков в прудах, в Чехии у молоди лосося на рыбоводном заводе, в России у молоди сиговых на рыбоводном заводе. Клинические признаки и патогенез. У рыб с большим числом каприниан изменяется окраска жабр. Так, у сиговых пораженные жабры бледно-розовые с желтовато-оранжевым оттенком. У американских сомиков отмечают нарушение целостности жабр, некроз отдельных участков, на которых часто поселяется сапролегния. Предполагается, что капринианы могут высасывать кровь из жаберных сосудов. 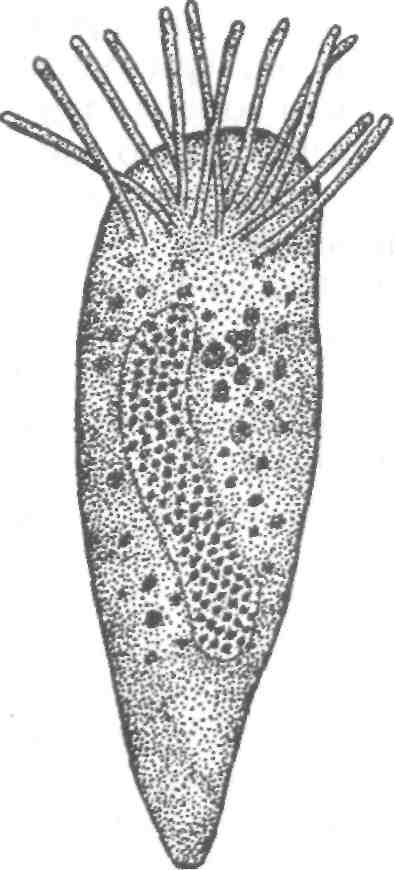 Рис. 61. Возбудитель каприниоза (трихофри-оза) Capriniana piscium Меры борьбы. Не разработаны. 8.2. БОЛЕЗНИ, ВЫЗЫВАЕМЫЕ КИШЕЧНОПОЛОСТНЫМИ 8.2.1. ПОРАЖЕНИЕ ИКРЫ ОСЕТРОВЫХ ПОЛИПОДИУМОМ Возбудитель. Polypodium hydriforme — единственный представитель кишечнополостных среди паразитов рыб. В большинстве случаев кишечнополостные — свободноживущие животные, часто колониальные, обитающие преимущественно в морях. Кишечнополостные — многоклеточные, двуслойные животные. В онтогенезе у них формируются только 2 зародышевых листка — экто- и эндодерма, отчетливо выраженные и у взрослых животных. В наиболее простом случае тело кишечнополостных представляет собой мешок. В полости мешка, выстланного эндодермой, происходит переваривание пищи. Отверстие мешка служит ртом, через него же выбрасываются непереваренные остатки пищи. Характерным признаком кишечнополостных является наличие в их покровах стрекательных клеток, служащих для прикрепления к субстрату, ловли добычи, защиты от врагов. P. hydriforme — паразит икры осетровых — имеет все признаки, характерные для кишечнополостных. Точное систематическое положение его не установлено. Развитие паразита происходит как в икре осетровых, так и в воде (рис. 62). Икра осетровых заражается паразитом в тот период, когда в ней начинается образование желтка (II—III стадия зрелости). В таких икринках находится самая ранняя из известных стадий развития — планулообразная двуслойная личинка. С ростом икринки и накоплением в ней желтка планула разрастается в длину и превращается в своеобразную трубку — столон, на котором образуются вздутия — почки, представляющие собой зачатки будущих самостоятельных особей. Число почек бывает различным. В мелкой икре севрюги их насчитывается 40—60, в более крупной икре осетра на столоне бывает 70—90 почек. В каждой почке столона закладывается по 12 щупальцев. К этому времени икра находится на IV стадии зрелости. Столон в этот период вывернут своим внутренним слоем (эндодермой) наружу и обращен к желтку, за счет которого происходит питание паразита. Перед нерестом при переходе яичников рыбы в V стадию зрелости столон с почками выворачивается так, что слои клеток принимают положение, характерное для кишечнополостных. Эндодерма оказывается обращенной во внутреннюю полость. При этом часть желтка икринки попадает в полость полипа. Во время нереста зараженные икринки выметываются вместе со здоровой икрой. Из них через разрыв оболочек столон выходит наружу и распадается на составляющие его почки, каждая из которых становится свободноживущим полипом. Свободноживущие полипы в течение первых 4—5 дней питаются за счет желтка, попавшего в их полость во время выворачивания столона. Затем у полипа образуется рот, и он переходит к питанию олигохетами, коловратками и другими мелкими водными животными, захватывая пищу своими щупальцами со стрекательными клетками. Полипы размножаются половым и бесполым путями. Бесполое размножение происходит путем продольного деления надвое в первую половину лета. С середины лета у полипов начинают образовываться половые железы. Мужские и женские половые железы чаще встречаются в разных особях, но имеются и гермафродит -ные полипы. Дальнейший ход развития паразита неизвестен. Неясно, как происходит оплодотворение и как паразит попадает в рыбу. Эти вопросы требуют дальнейшего изучения. 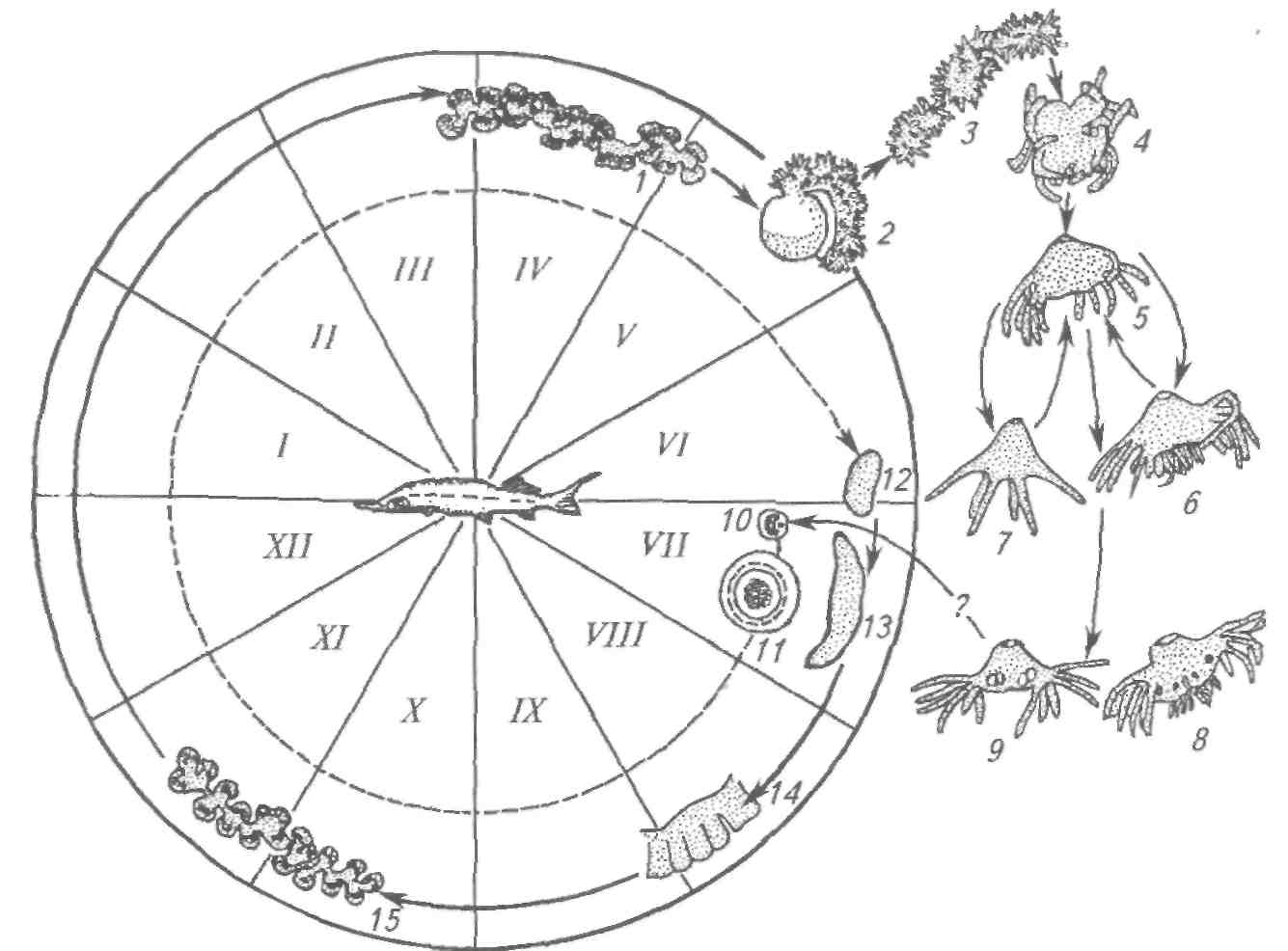 Рис. 62. Цикл развития Polypodium (внутри круга помещены паразитические стадии развития — внутри ооцита); вне круга — свободно-живущие; римскими цифрами обозначены месяцы: 7, 15 — столон с почками, щупальцами внутри почек; 2 —столон с наружными щупальцами, выходящий в воду во время нереста; 3 —такой же столон в воде; 4 —фрагмент столона в воде; 5 — одиночная медуза с щупальцами; 6— 24-щупальцевая медуза; 7—6-щупальцевая медуза; 8— 24-щупальцевая медуза с незрелыми половыми железами первого типа; 9—12-щупальцевая медуза со зрелыми половыми железами второго типа; 10 — двухъядерная клетка; 11 — морула; 12— планула; 13— почкующаяся планула; 14 — столон с почками без щупальцев. Знак ? означает, что способ заражения ооцитов неясен Эпизоотология. Полиподиум отмечен практически во всех водоемах, где обитают осетровые. Он паразитирует в икре стерляди, осетра, севрюги, шипа, белуги, калуги и веслоноса. Имеются сведения о нахождении его у американских осетровых. Клинические признаки и патогенез. По внешнему виду рыбы не видно, заражена икра или нет. Однако зараженные икринки отличаются от незараженных более крупными размерами и цветом. На первых стадиях развития они темнее незараженных, на более поздних — светлее. Пораженные икринки погибают. Паразитирующий в икре полиподиум снижает репродуктивную способность зараженных рыб и тем самым влияет на численность воспроизводимого стада. Кроме того, из-за зараженности икры снижается ее пищевая ценность. Диагноз. Его ставят на основании нахождения паразита в икре осетровых. Зараженность определяют при вскрытии рыб или пробу берут из ястыка живой рыбы с помощью специального щупа. В самках рыб II степени зрелости, а также в ооцитах младшей генерации зараженные икринки внешне не отличаются от зрелых, и диагноз ставят на основании исследования гистологических срезов. Меры борьбы. Водоем или рыбоводное хозяйство, в которых обнаружена инвазированная P. hydriforme рыба, объявляется неблагополучным и на него накладываются ограничения. Лечение зараженных полиподиумом осетров не разработано. Применяют различные профилактические мероприятия для предотвращения расселения паразита и снижения его численности в водоеме. Достигается это строгим контролем за перевозками. Перевозить осетровых можно только из благополучных водоемов и на стадии оплодотворенной икры. Нельзя перевозить созревших рыб даже на ранних стадиях зрелости. Необходимо также следить за тем, чтобы на предприятиях икорного производства зараженная икра не попадала в воду. Поэтому отходы икорного производства следует собирать в строго отведенные для этого места, обезвреживать и утилизировать. 8.3. ГЕЛЬМИНТОЗЫ Гельминтозами (от греч. helmintos — червь) называют заболевания, вызываемые паразитическими червями (гельминтами). Классификация гельминтов постоянно меняется. Согласно одной из них, изложенной в многотомном издании «Определителя» (1985*, 1987**), гельминты рыб относятся к четырем типам животного царства: плоским червям (Plathelminthes), нематгельминтам (Nemathelminthes), скребням (Acanthocephales) и кольчатым червям (Annelida). При изложении материала мы в основном использовали этот «Определитель», однако в некоторых случаях внесены правки с учетом последних мировых разработок. Из типа плоских червей заболевания у рыб вызывают в основном представители классов: Monogenoidea, Amphilinida, Cestoda, Trematoda. 8.3.1. МОНОГЕНОИДОЗЫ Возбудители моногеноидозов — представители класса моногеней Monogenea. Моногеней относятся к плоским паразитическим червям, имеют плоское, сплющенное в спинно-брюшном направлении тело. Размеры моногеней значительно варьируют. Длина тела колеблется от долей миллиметра до нескольких сантиметров. На переднем конце расположены 2—6 лопастей, на которых открываются протоки головных желез. Реже имеется только одна лопасть. На лопастях могут быть различные прикрепительные образования: ямки, валики, присоски. Задний конец представляет собой прикрепительный диск, снабженный различными, иногда довольно сложными крючками, клапанами, присосками. В отдельных случаях прикрепительный диск превращается в одну большую присоску. Число и форма прикрепительных образований служат основой для определения вида моногеней. Покровы моногеней представлены погруженным эпителием, затем следуют кольцевые слои: мускулатура, диагональные и продольные пучки мышц, с помощью которых осуществляется движение прикрепительных образований. Нередко покровы образуют кольцевую складчатость, имеют кутикулярные шипы для лучшего прикрепления к хозяину. Полость тела заполнена паренхимой, в которой располагаются внутренние органы. Пищеварительная система начинается на переднем конце тела субтерминально расположенным ротовым отверстием, окруженным околоротовой воронкой. Ротовое отверстие переходит в окологлоточную сумку, в которую вдается мускулистая глотка. За глоткой следует пищевод, причем он может отсутствовать. Далее следует кишечник, состоящий из одного или двух стволов. Кишечные стволы могут быть гладкими или иметь выросты, часто сливающиеся друг с другом и образующие своеобразную сетку. Стволы могут либо заканчиваться слепо, либо сливаться между собой, образуя кольцо. * Определитель паразитов пресноводных рыб фауны СССР. Т. 2. Паразитические многоклеточные (первая часть). — Л.: Наука, 1985. — 425 с. ** Определитель паразитов пресноводных рыб фауны СССР. Т. 3. Паразитические многоклеточные (вторая часть). — Л.: Наука, 1987. — 583 с. Нервная система состоит из парного надглоточного ганглия и отходящих от него нервных стволов. У многих моногеней имеются глаза, число и расположение которых могут варьировать. Выделительная система представлена протонефридиями и системой длинных каналов, открывающихся двумя отверстиями на уровне глотки. Моногеней — гермафродиты. У них имеется один, два или много семенников, расположенных обычно в заднем конце тела. От семенника отходит семяпровод, переходящий в семяизвергатель-ный канал, открывающийся через копулятивныи орган в половой атриум. Копулятивныи орган представляет собой или мускулистый пенис, или хитиноидное образоване, состоящее из хитиноид-ной трубки с поддерживающим аппаратом. Форма и размеры ко-пулятивного органа служат систематическим признаком при определении видов. Яичник один, причем расположен перед семенниками. В короткий яйцевод впадают протоки желточников, фолликулы которых расположены по бокам тела от глотки и до прикрепительного диска. Яйцевод переходит в оотип. В нем формируются яйца, часто снабженные ножками и филементами, с помощью которых они прикрепляются к подводным предметам или редко к поверхности тела рыб. У некоторых моногеней (гиродактилусы) яиц не образуется и они отрождают живых особей, почти полностью сформировавшихся червей. Для этой группы моногеней характерно формирование нескольких поколений гельминтов в еще не родившихся особях. У яйцекладущих форм яйца попадают в воду, опускаются на дно или прикрепляются к различным подводным предметам и растениям. Реже они прикрепляются к жабрам или поверхности тела хозяев. Для достижения инвазионности отложенные яйца должны пройти процесс развития. Срок развития яиц зависит от температуры и может растягиваться на очень длительные сроки. Так, яйца Dactylogyrus vastator, отложенные осенью, остаются живыми до следующей весны. Из созревшего яйца вылупляется личинка (он-комирацидий), которая плавает в воде с помощью имеющихся на ее поверхности ресничек. Плавающая в воде личинка встречает рыбу—хозяина, прикрепляется к ней, сбрасывает ресничный покров и начинает паразитический образ жизни. Моногеней не всегда сразу попадают к своему постоянному месту обитания. Иногда им приходится переползать с одной части рыбы на постоянное место паразитирования. Например, представители рода Dactylogyrus паразитируют на поверхности тела и плавниках мальков, пока у последних не сформировались жабры, и переползают на жабры только после того, как они полностью сформируются. Моногенеи обладают, как правило, строгой специфичностью и паразитируют обычно в одном, реже в нескольких близкородственных хозяевах. Большинство моногенеи — эктопаразитические животные, живущие на поверхности тела и жабрах рыб. Среди моногенеи много возбудителей опасных болезней, приводящих к гибели рыб. 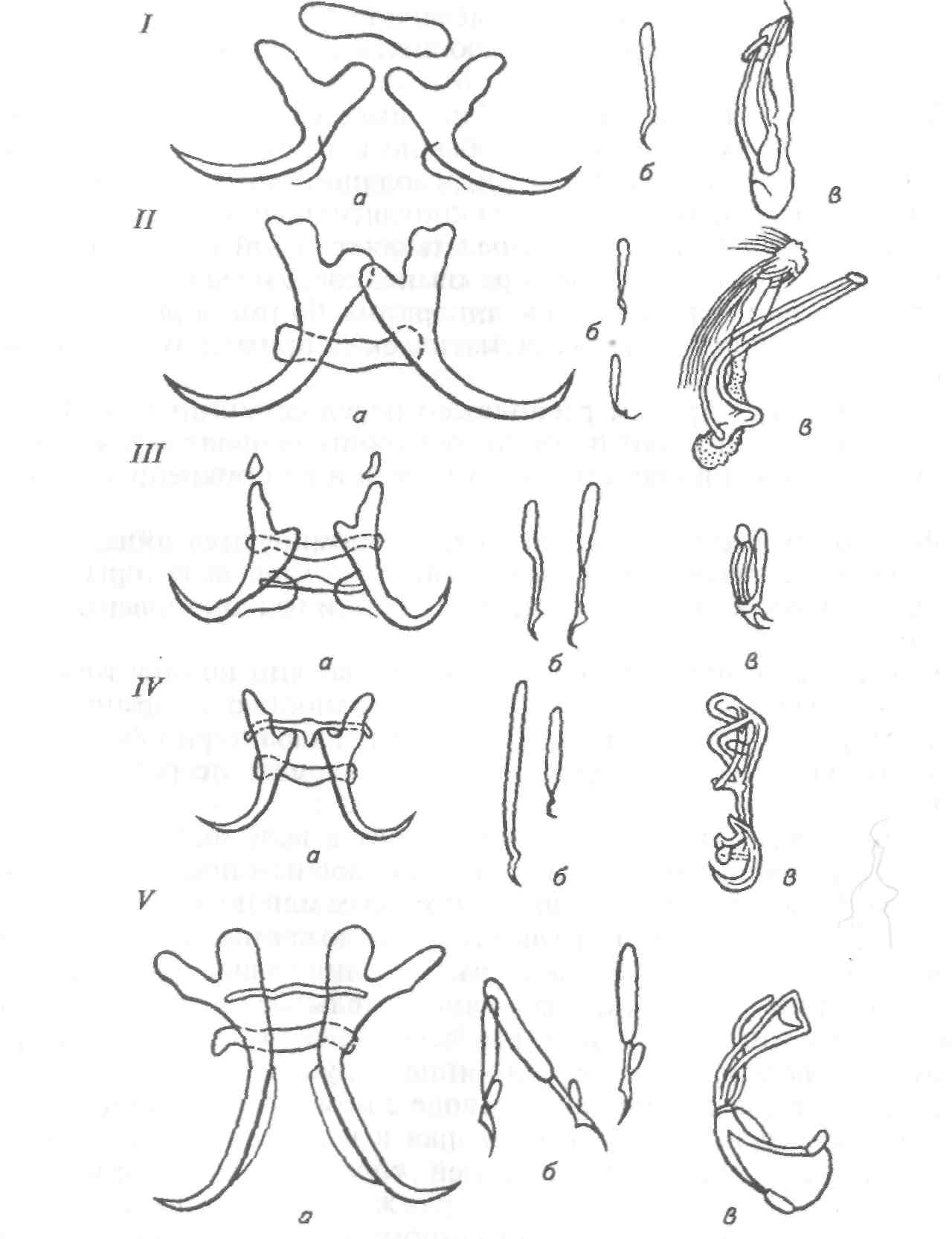 Рис. 63. Прикрепительные крючья и копулятивные органы дактилогирусов: / — Dactylogyrus vastator; // — D. extensus; III — D. lamellatus; IV— D. hypophthalmichthys; V— D. aristichthys; a — серединные крючья; б — краевые крючья; в — копулятивный орган Класс моногенеи Monogenea делится на 2 подкласса: низшие моногенеи Polyonchoinea и высшие моногенеи Oligonchoinea. Низшие моногенеи чаще встречаются на пресноводных, высшие — на морских рыбах. Дактилогирозы карпа Возбудители дактилогирозов карпа относятся к сем. Dactylogyridae. Для них характерны небольшие размеры тела (не более 2 мм), наличие глаз на его переднем конце. В состав прикрепительного диска входят 14 краевых крючьев, 2 средних крючка и 1 соединительная пластина. Кишечник двуветвистый, чаще без выростов, заканчивается слепо или образует кольцо. Представители семейства — яйцекладущие организмы. Яичник округлый, вытянутый или колбасо-видный. Вагинальный проток, как правило, одинарный, открывается на боку тела, часто снабжен хитинодной выстилкой. Жел-точники сильно развиты, парные, расположены по бокам тела. Семенник, как правило, один, причем расположен позади яичника. От семенника отходит семяпровод, заканчивающийся копуля-тивным органом иногда довольно сложного строения. Яйца у разных видов различаются по форме. Форма и структура прикрепительных крючьев и копулятивного органа служат признаками для определения вида. Представители семейства — узкоспецифичные паразиты — обитают чаще на жабрах, редко на поверхности тела и в носовых ямках. На жабрах карпов, сазанов и их гибридов паразитируют 5 видов дактилогирусов, однако эпизоотическое значение имеют только Dactylogyrus vastator и D. extensus (рис. 63). |