ихтиопаталогия. Дана характеристика рыб как переносчиков возбудителей болезней человека и животных. Для студентов вузов по специальности Водные биоресурсы и аквакультура
 Скачать 10.48 Mb. Скачать 10.48 Mb.
|

|
Тетраонхоз сиговых Возбудитель. Моногенетический сосальщик Tetraonchus alasken-sis из сем. Tetraonchidae, паразитирующий на жабрах сиговых. Черви средних размеров (длина тела до 2,3 мм, ширина — 0,6 мм). Прикрепительный аппарат с 16 краевыми и двумя парами серединных крючьев и одной соединительной пластиной. Кишечник в виде одиночного ствола без боковых выростов. Развитие прямое. Паразит размножается яйцами. Возбудитель обнаружен у сиговых в бассейне р. Обь. Паразиты встречаются у пыжьяна, чира, пеляди и нельмы. Сильнее других заражается пелядь, что ведет к ее гибели. Наибольшая зараженность и гибель пеляди отмечаются у старших возрастных групп с августа до ледостава. У сильно зараженных рыб отмечают участки с многочисленными кровоизлияниями и некрозом жабр. В отдельных случаях наблюдается вторичное поражение сапролегнией. Диагноз. Его ставят по клиническим признакам и при обнаружении большого числа возбудителей. Меры борьбы. Регулирование численности стада пеляди. Лечение не разработано. Из других представителей этого рода широко распространен Т. monenteron, локализующийся на жабрах щук (рис. 68). 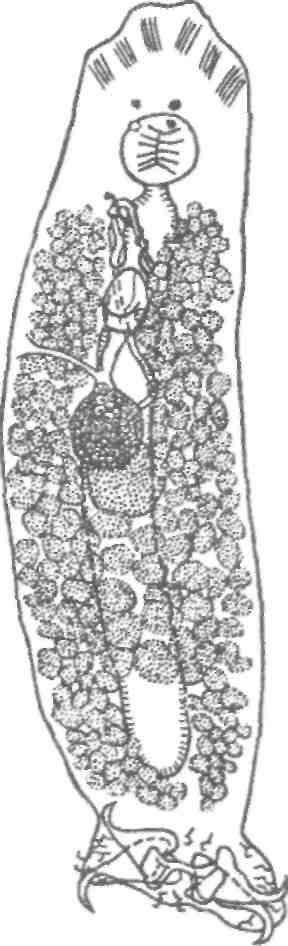 Рис. 68. Tetraonchus monenteron Другие моногенеи пресноводных рыб Наряду с моногенеями, о которых известно, что они вызывают заболевания, на рыбах встречаются виды, которые можно отнести к потенциально опасным. Среди них в первую очередь следует отметить моногенеи из рода Diplozoon сем. Diplozoidae (рис. 69). Это очень своеобразные черви, во взрослом состоянии представляющие собой 2 накрест сросшиеся особи (спайник или дипорпа). Прикрепительный диск несет 2 пары серединных крючьев и 4 пары прикрепительных клапанов. На рыбах встречается несколько видов этого рода. Длина тела каждой сросшейся особи достигает 4—6 мм. На жабрах рыб спайники встречаются иногда в значительных количествах как в естественных водоемах, так и в прудовых хозяйствах. Заболеваний, вызываемых диплозонами, пока отмечено немного. На жабрах осетровых в различных водоемах паразитируют моногенеи из рода Diclybothrium сем. Diclybothriidae (рис. 70). Это довольно крупные черви длиной 4—23 мм. Тело листовидное, вытянутое, на заднем конце располагается хорошо отграниченный от тела прикрепительный диск, несущий 3 пары прикрепительных клапанов, внутри которых расположены крупные крючки. На заднем конце прикрепительного диска имеется вырост, несущий 3 пары крупных и 1 пару очень мелких крючков, которые трудно рассмотреть. Заболеваний этот паразит пока не вызывал. При культивировании угря могут встречаться Gyrodactylus anguillae и два вида моногеней рода Pseudodactylogyrus: P. bini и P. microrchis. Обычно они поражают молодь угря в первый год выращивания. Описаны в угревых хозяйствах Японии и в нашей стране. Все паразиты строго специфичны по отношению к хозяину, теплолюбивы и могут вызывать заболевание и высокую гибель рыб. Для борьбы с паразитами эффективны аммиачные ванны. 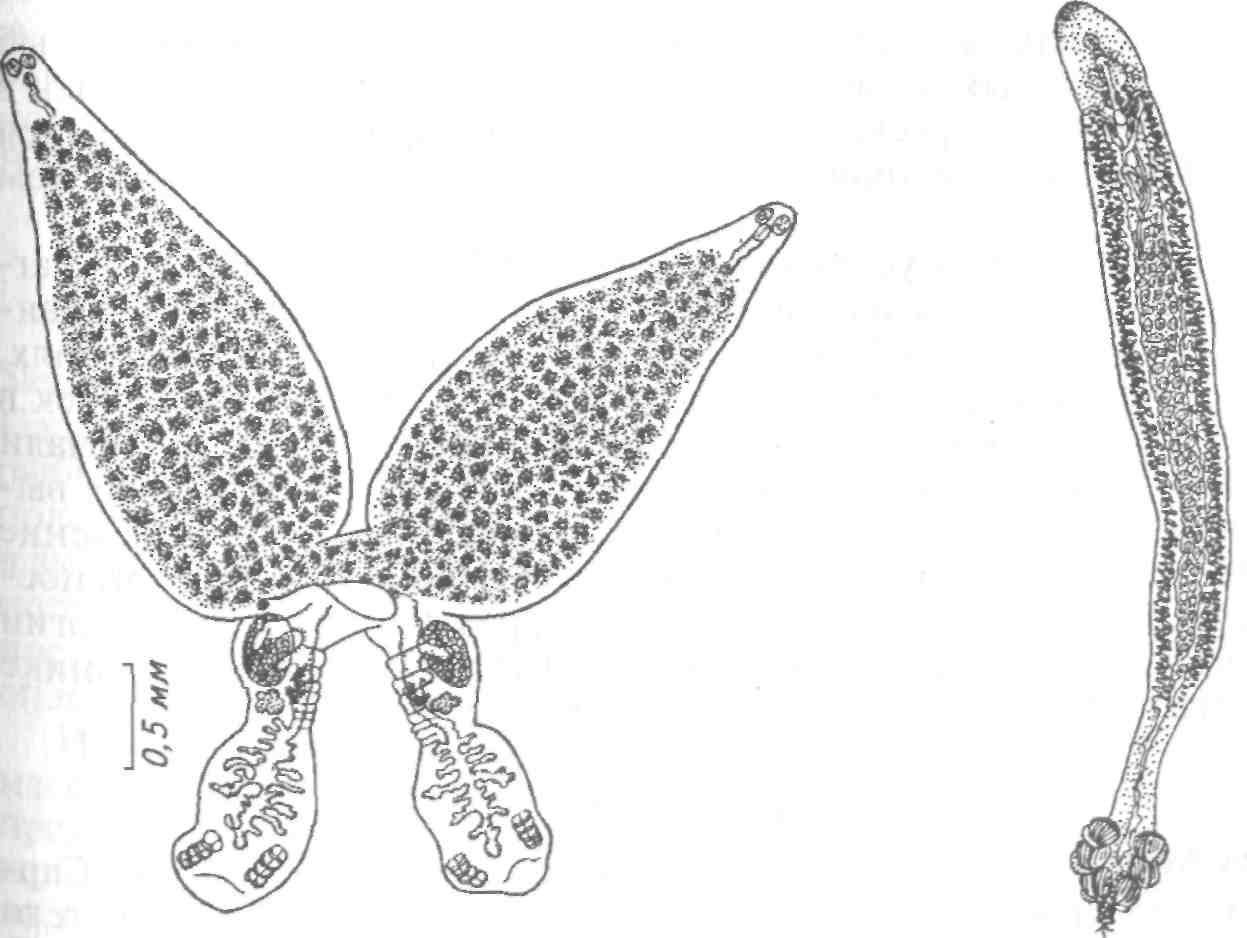 Рис. 69. Diplozoon paradoxum Рис 70. Diclybothrium armatum Моногеней морских рыб Моногеней широко представлены у морских рыб, заражая в основном жабры, носовые ямки, поверхность тела, плавники. Иногда они встречаются в глотке, мочеточниках, клоаке хрящевых рыб, и, как исключение, в кровеносной системе рыб. При обилии видов моногеней у морских рыб вероятность заражения ими рыб в естественных водоемах невелика и они не представляют угрозы для хозяина. Патогенного воздействия не обнаруживается даже при значительной экстенсивности инвазии. Однако моногеней могут стать серьезной угрозой при выращивании морских рыб в аквариумах и хозяйствах марикультуры. Они относятся к разным семействам. Среди них встречаются довольно крупные формы, размеры которых превышают 1,5—2,5 см. Гиродактилусы морских рыб Гиродактилусы довольно часто встречаются у морских рыб иногда в массовых количествах. Так, Gyrodactylus marmus найден в Тихом океане на треске (в тысячах экземпляров). Он встречается также в Белом, Баренцевом, Норвежском, Охотском и Японском морях. G. arcuatus и G. bychowskyi паразитируют на колюшках. G. аг-cuatus локализуется на жабрах и плавниках трехиглой и девя-тииглой колюшек в Белом, Баренцевом и Балтийском морях. G. bychowskyi найден на жабрах трехиглой и амурской колюшек в Белом, Баренцевом, Балтийском и Японском морях. Отмечали случаи массового заражения и гибели колюшек в Белом море, вызванные данными видами гиродактилусов. Массовое заражение зеленушек (Grenilabrus griseus) вызывал Gyrodactylus crenilabri после пересадки их из Черного моря в аквариум Института биологии южных морей. Через 7—10 дней у зеленушек на каждом плавнике насчитывали от 90 до 100 паразитов и более. Бенедениоз Возбудитель — Benedenia seriola (рис. 71) относится к сем. Сар-salidae. У червей длиной 5,5—6,6 мм и шириной 3,1—3,9 мм тело удлиненно-овальное, плоское. На переднем конце тела расположены эллипсовидные головные присоски. Прикрепленный диск крупный, с зазубринами по краям. Он разделен по внутренней поверхности на центральный и семь краевых участков, по краю которых расположены 14 очень маленьких крючков и три пары срединных, причем последние лежат попарно друг за другом в центре задней части прикрепительного диска. Кишечник ветвистый. В. seriola локализуется на коже желтохвоста (Seriola quinquera-diata) из сем. ставридовые. Развитие прямое. Паразит неоднократно продуцирует яйца, которые попадают в воду. Выклюнувшаяся из яйца личинка может жить без хозяина в течение дня. Развитие от вылупления личинок до созревания червей длится не менее 18 дней при температуре 22—26 0С. В. seriola поражает желтохвоста, разводимого в закрытых лагунах Японии. На одной рыбе находили до 570 крупных паразитов. Зараженность желтохвостов изменяется по сезонам. Наиболее низкая инвазия наблюдается в зимний и весенний периоды. При сильной инвазии у желтохвоста снижается темп роста, рыба истощается, уменьшается плодовитость. Больные рыбы чаще подвергаются бактериальному дерматиту. В марикультуре при поражении мо-ногенеями применяют лечебные ванны. При поражении рыбы В. seriola используют ванны из пресной воды и пирофосфата натрия. 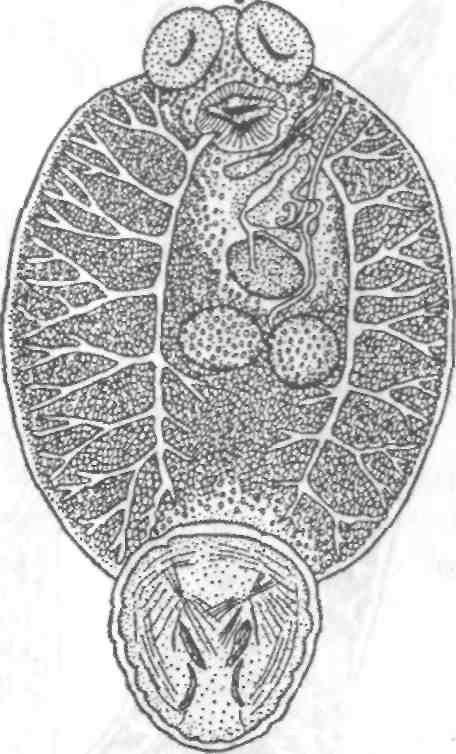 Рис. 71. Benedenia seriola Другие моногенеи морских рыб В морских аквариумах наблюдались эпизоотии, вызванные Neobenedenia melleni — червями длиной 5 мм и шириной 3 мм. Паразит поражал каменных и рифовых окуней. С зараженной рыбы снимали до 2000 червей, заселявших глаза, жабры и носовую полость. При таком заражении рыба погибает. N. melleni поражает в аквариумах 48 видов рыб из 17 семейств и трех отрядов. Для моногенеи это редкое явление. В аквариумах не исключена возможность более широкого заражения, чем в природе. Представляет опасность для тиляпии, выращиваемой в марикультуре. Известен целый ряд моногенеи, которые становятся причиной массовой гибели рыб в марихозяйствах. К их числу относятся, прежде всего, представители семейств диплектанид, микрокотилид, Капсалид и некоторые другие. Так, моногенея Allobivagina sp. вызвала гибель рыбы в марихозяйствах, размещенных на Красном море. Длина гельминта 40—80 мм. Количество паразитов на рыбах достигало 50—1133 экз. У пораженных рыб появлялись признаки анемии, разрушались жаберные лепестки, наблюдалось усиленное выделение слизи, что затрудняло дыхание. При разведении желтохвоста на жабрах встречается моногенея Не-teraxine heterocerca (рис. 72) из сем. Axinidae, длиной 10—20 мм. На прикрепительном диске 23—33 клапана на длинной стороне и по 5— 10 клапанов на короткой стороне. Высокая зараженность может вызвать острую анемию и гибель хозяина. Даже незначительное количество паразитов вызывает истощение рыб. Тяжелое заражение желтохвоста этой моногенеей наблюдалось в ряде районов Японии. Bivaginia tai (сем. Microcotylidae) вызывает серьезные заболевания жабр красного тая (Pagrus major), успешно разводимого в Японии. Длина червей 7— 9 мм, ширина 0,54 мм. На прикрепленном диске по 38—60 клапанов с каждой стороны. Гибель рыб в морском аквариуме вызывает Calceostomella inermis (рис. 73) из сем. Calceostomatidae. Это черви длиной 5— 9 мм, шириной 0,5—1,0 мм. Паразитируют на жабрах, плавниках, коже и глазах темного горбыля в Черном и Средиземном морях. В Севастопольском аквариуме иногда наблюдается размножение С. inermis в огромных количествах. Черви покрывают всю поверхность тела рыб сероватым налетом. На рыбе находили по 400—1000 экз. паразита. При поражении кожи горбылей несколькими сотнями экземпляров С. inermis отмечена гибель рыб. Возникновение эпизоотии в аквариуме можно объяснить ослабленно-стью рыбы и тесным контактом между горбылями, а также и более благоприятными условиями для развития личинок гельминта. Microcotylemugilis — обычный паразит кефалей, широко распространенный на жабрах лобана, головача, сингиля и остроноса в Каспийском, Черном и Средиземном морях. Длина червей 4,8—7,3 мм, ширина 1,3—1,52 мм. На прикрепительном диске располагается до 30 пар клапанов. Axinebelone — паразит сарганов. Род Axine близок к роду Microcotyle, но отличается от него асимметричным диском, расположенным косо по отношению к продольной оси тела. Длина A. belone 4—6 мм, ширина 1,0—1,5 мм. Число клапанов зависит от возраста червя. Кроме сарганов, A. belone встречается у ставридовых. Переход паразитов с одной группы хозяев на другую связан с наличием экологического контакта между ними, а не их генетической близостью. Mazocraes alosae (сем. Маzocraeidae) паразитирует на жабрах сельдей рода Alosa в бассейнах Каспийского, Черного, Азовского, Средиземного, Северного морей и в Южной Атлантике. Длина М. alosae (рис.74) 2—12мм, ширина 0,5—1,7 мм. Прикрепительный диск ромбической формы, имеет по 4 канала с каждой стороны. Паразитирует у рыб старших возрастов. Кладка яиц начинается в Каспии во второй половине мая, причем яйца прикрепляются на том же хозяине к поверхности тела, жабрам, внутренней поверхности жаберной крышки. Через 8—10 дней после начала кладки из яйца выходят личинки, которые локализуются на жабрах хозяина. 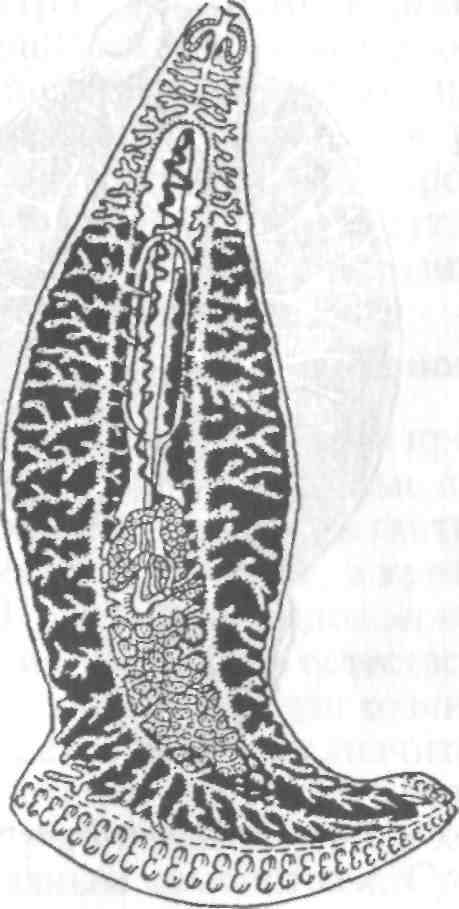 Рис. 72. Heteraxine heterocerca Размножение М. alosae длится около 1 мес. Весь этот период полностью совпадает со сроком икрометания сельдей, которые подходят к берегам или заходят в реки на нерест, образуя при этом большие концентрации. В таких условиях и происходит перекрестное заражение рыб. Короткий срок размножения червя и закрепление им яиц на хозяине являются приспособлением, обеспечивающим существование вида. Сельди младших возрастов не подходят к берегам, не контактируют с сельдями старших возрастов, поэтому свободны от паразита. 8.3.2. АМФИЛИНОЗ Amphilina foliacea и A. japonica — представители особого класса — амфилин (Amphilinida), сем Amphilinidae, паразитирующие в полости тела осетровых рыб. Возбудители. Половозрелые гельминты листовидно-овальной формы, белого или слегка кремового цвета, достигающие максимального размера 26—124x12—42 мм (рис. 75, 76). Тело их не расчленено и имеет характерную ячеистую поверхность. Передний конец снабжен втяжным хоботком. Матка трубкообразная, расположенная по всему телу в виде горизонтальных и вертикальных петель, открывается рядом с хоботком. Многочисленные округлые семенники лежат между петлями матки. Яичник рассеченный, желточники тянутся вдоль боковых сторон тела. У половозрелых особей матка заполнена яйцами. По мере продвижения яиц по ветвям матки в них происходит формирование ресничной, вооруженной 10 крючочками эмбриона ликофоры. Известно всего два вида амфилин. В России обнаружены оба. 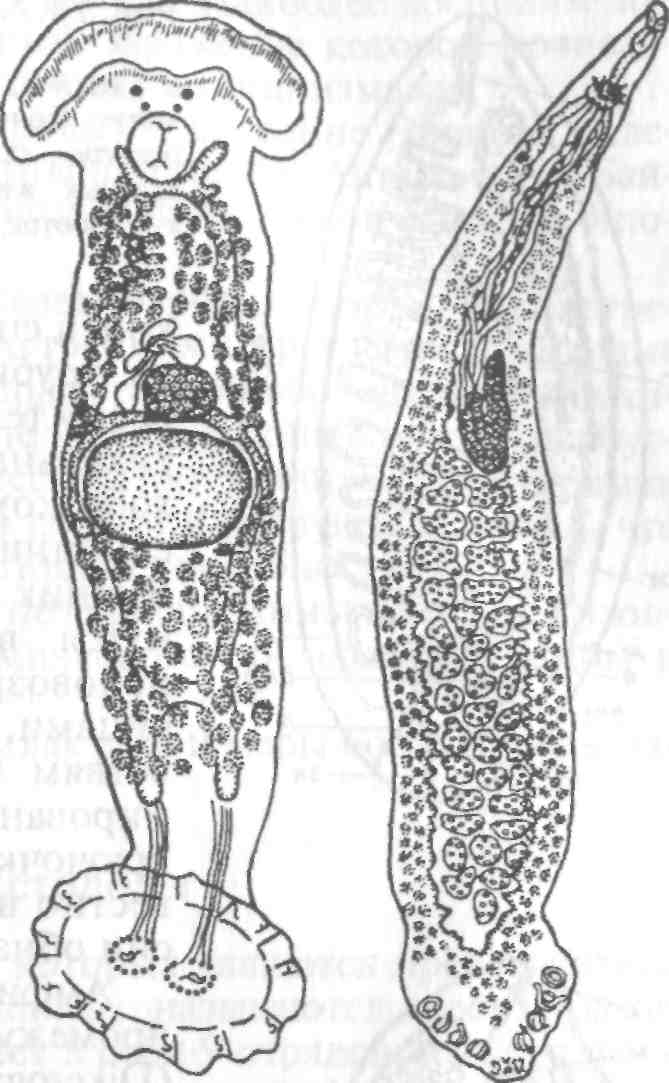 Рис. 73. Calceostomella inermis Рис. 74. Mazocraes alosae 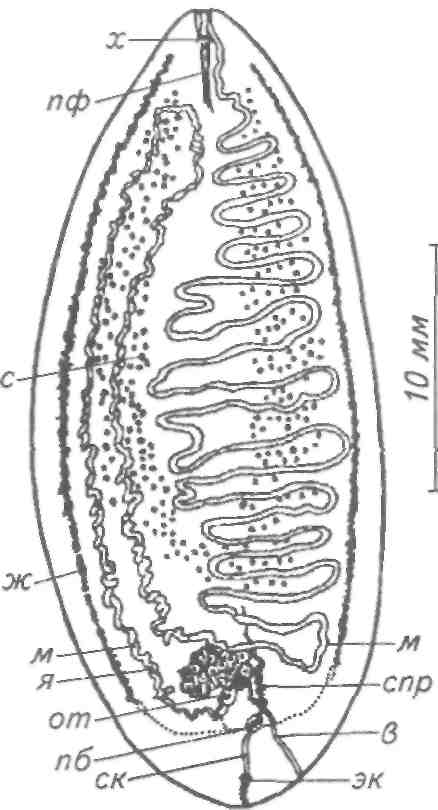 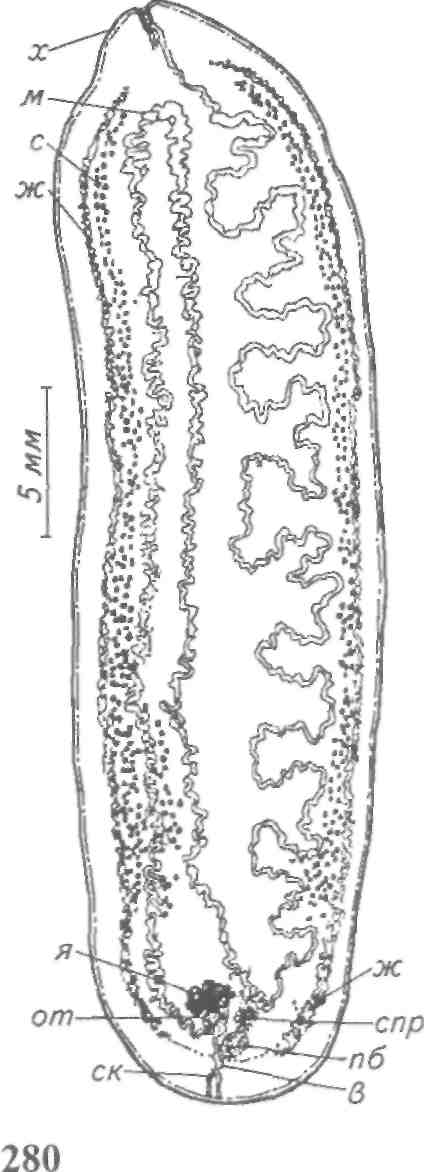 Рис. 75. Amphilina foliacea (общий вид с брюшной стороны): в —вагина; ж —желточные железы; м — матка; от — оотип (тельце Мелиса); пб — пропульсаторный буль-бус; пж — простатические железы; пф — протоки фронтальных желез; с — семенники; спр — семяприемник; х —хоботок; эк— эмбриональные крючья; я —яичник Амфилины развиваются с участием промежуточных хозяев — бокоплавов (Dikerogammarus caspius, D. haemobaphes, Pontogammarus crassus, P. obesus и др.). В полости тела бокоплавов ликофора претерпевает метаморфоз и превращается в высокоорганизованную личинку, обладающую всеми системами органов, свойственными взрослым червям. Осетровые рыбы заражаются при поедании инвазированных бокоплавов. При помощи хоботка личинки проникают сквозь стенки желудка в полость тела рыбы, где растут, заканчивают половое созревание и продуцируют яйца. Зрелые яйца через абдоминальную пору рыб попадают в воду. Эпизоотология. A. foliacea паразитирует у белуги, шипа, севрюги, русского, сибирского и атлантического осетров и стерляди. A. japonica выявлена в полости тела сахалинского и амурского осетров и калуги. Наиболее восприимчива к заражению молодь осетровых рыб, заражение которой происходит при переходе на корм планктонными организмами. Рис. 76. Amphilina japonica (общий вид с брюшной стороны) условные обозначения те же, что на рис. 75 A. foliacea встречается у осетровых в бассейне Черного, Каспийского морей и реках Обь, Иртыш, Енисей, Ангара и оз. Байкал. A. japonica — паразит осетровых рыб Амурской области, Японии и Северной Америки. Клинические признаки и патогенез. Взрослые амфилины встречаются в полости тела между внутренними органами, а молодые особи также и в тканях печени, под серозной оболочкой. Они оказывают механическое воздействие на окружающие органы и обусловливают атрофические процессы. При значительной инвазии отмечены снижение гликогена в печени и разрушение гонад, что приводит к снижению репродуктивной способности рыб. Диагноз. Для постановки диагноза необходимы данные эпизоо-тологического и патологоанатомического обследования рыбы и обнаружение амфилин. Меры борьбы. Лечение, профилактика и меры борьбы не разработаны. 8.3.3. ЦЕСТОДОЗЫ Гельминтозы, возбудителями которых являются представители класса ленточных червей Cestoidea, называются цестодозами. Класс ленточных червей включает в себя 9 отрядов, из которых в костистых пресноводных рыбах на взрослой фазе встречаются представители четырех отрядов: Caryophyllidea, Pseudophyllidea, Proteocephalidea и Nippotaeniidea. Представители четырех других отрядов (Trypanorhyncha, Diphyllidea, Tetraphyllidea и Lecanicepha-lidea) во взрослом состоянии паразитируют в акулах и скатах, а в костистых морских и проходных рыбах встречаются только их личинки — плероцеркоиды. Девятый отряд — Cyclophyllidea — объединяет высших цестод—паразитов теплокровных животных, у которых личиночные формы отдельных видов могут развиваться в различных внутренних органах пресноводных рыб. Среди ленточных червей далеко не все виды вызывают заболевания рыб. Однако есть виды весьма патогенные, вызывающие значительные эпизоотии, сопровождающиеся гибелью рыбы. Тело ленточных червей молочно-белого цвета, плоское, лентовидное, состоит из головки (сколекса) и множества члеников (проглоттид), составляющих стробилу. Число члеников может достигать сотен и даже тысяч. У некоторых представителей этого класса, в частности отряда Caryophyllidea, тело не имеет члеников и выглядит как сплошная лента. Длина тела некоторых цестод может достигать 10—15 м. Половозрелые ленточные черви чаще всего паразитируют в кишечнике, личиночные стадии (плероцеркоиды) — в полости тела и других органах и тканях рыб. Сколекс цестод служит для прикрепления и потому снабжен различными органами, приспособленными для этого: ботриями, ботридиями, присосками, хоботками. Прикрепительные органы надежно удерживают гельминта в кишечнике (рис. 77). Наиболее примитивными органами прикрепления являются ботридии, которые представляют собой присасывательные ямки и бороздки на головке (1—4 у гвоздичников). Ботридии более разнообразны. Это — хорошо обособленные прикрепительные органы с развитой мускулатурой, располагающиеся на брюшной или спинной стороне сколекса — округлые, полые образования, снабженные собственной сильной мускулатурой, например у протеоцефалюсов. Хоботки — мышечные выросты, расположенные в верхней части головки; они могут втягиваться в специальное хоботное влагалище (четыреххоботники); хоботки обычно вооружены крючьями. Помимо хоботка хитиноидные крючья разнообразной формы и величины могут быть расположены на ботридиях (триенофорусы). За головкой следует шейка — зона роста, где формируются членики. Размер шейки различен. Молодые членики находятся сразу же за шейкой, старые — по мере роста червя отодвигаются все дальше от нее. В каждом членике имеется набор половых органов, которые развиваются в определенной последовательности: сначала закладываются элементы мужской половой системы, позднее женской, и членики становятся гермафродитнами. Последние членики стробилы почти целиком заполнены маткой, набитой яйцами. Тело ленточных червей покрыто кутикулой, под которой находится кожно-мускульный слой. Полости тела у них нет и пространство между внутренними органами заполнено губчатой паренхиматозной тканью. Под кожно-мускульным слоем расположены продольные и поперечные мышцы. Нервная система цестод представляет собой центральное нервное кольцо, располагающееся в головке. От него вперед и назад отходят продольные, нервные стволы, которые в каждом членике соединяются между собой поперечными кольцевыми комиссурами. 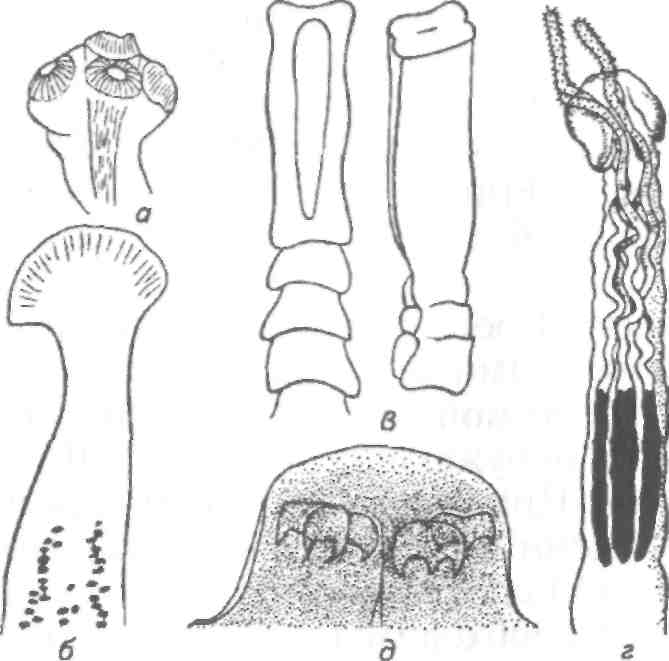 Рис. 77. Органы прикрепления цестод: а —присоски Silurotaenia siluri; б —ботрии Caryophyllaeus laticeps; в — ботридии Bothriocephalus scorpii; г — хоботок Tetrarhynchus; д— крючья Triaenophorus meridionalis червя (ботриоцефалюс). Присоски Выделительная система состоит из мерцательных клеток, располагающихся в паренхиме. От них отходят мелкие канальцы, которые соединяются с выводными протоками (их 2 или 4). Они проходят в боковых частях члеников (соединяясь между собой комиссурами), в конце стробилы объединяются в экскреторный пузырек, открывающийся наружу. У нечленистых червей (гвоздичников) и ремнецов поперечные комиссуры отсутствуют и продольные выделительные протоки соединяются между собой беспорядочной сосудистой сетью. Питание червей происходит путем всасывания всей поверхностью тела через кутикулу, на которой имеются микроскопические микротрихии, имеющие сложное строение, отличное от микро-ворсинок других плоских червей. Половая система у расчлененных цестод самостоятельная в каждом членике, у нерасчлененных цестод имеется всего один половой комплекс. Все цестоды рыб — гермафродиты. Мужской половой аппарат состоит из семенников и выводных путей. Количество семенников различно: чаще их десятки, даже сотни, реже единицы. Они расположены в паренхиме, ближе к спинной стороне тела. От семенников отходят тонкие семявыносящие канальцы, которые сливаются в семяпровод, причем последний на конце может преобразовываться в семенной пузырек. Семяпровод идет к копулятивному органу — циррусу, способному выпячиваться; циррус расположен в мышечном мешке, называемом сумкой цир-руса. Входящая в сумку цирруса часть семяпровода является семя-извергательным каналом. Циррус и влагалище открываются в половую клоаку. Женская половая система состоит из яичников, желточников, дополнительных желез, протоков и резервуаров, служащих для хранения и выведения половых продуктов. В каждом половом комплексе имеется один, чаще двухлопастной яичник, от которого отходит яйцевод. В него открывается семяприемник и далее общий проток ведет к оотипу, в который поступают протоки желточников и железы Мелиса, способствующей образованию скорлуповой оболочки. В оотипе формируются яйца. Желточники чаще состоят из многочисленных фолликулов, которые располагаются в паренхиме, либо вокруг внутренних органов, либо в боковых частях члеников. От желточников отходят мелкие и тонкие желточные протоки, сливающиеся в два крупных протока, которые далее объединяются в один и открываются в оотип. Вагина представляет собой трубку, связывающую женское половое отверстие с семяприемником, находящимся около оотипа. От оотипа берет начало матка, которая у многих цестод рыб имеет вид сильно извитой трубки, иногда мешковидной. Зрелые яйца выходят из матки в воду через ее отверстие, открывающееся самостоятельно на спинной или брюшной стороне членика или нерасчлененного червя. Помимо этого, зрелые яйца могут попадать в воду вместе с отторгнутыми члениками стробилы. При совокуплении цестод циррус выпячивается и проникает в вагину этого же или соседнего членика или членика другой стробилы, лежащей рядом. Соответственно происходит самооплодотворение или перекрестное оплодотворение. Эмбриональное развитие цестод рыб происходит в яйце, находящемся в большинстве случаев в матке, и попадающие в воду зрелые яйца содержат сформированных эмбрионов, имеющих 6 крючьев (онкосфера) или 10 крючьев (ликофора). В воде из яйца выходит личинка, называемая корацидием, которая некоторое время ведет свободный образ жизни. Тело корацидия покрыто ресничками, имеет 3 пары подвижных эмбриональных крючьев. У некоторых цестод личинки не выходят из яйца, а попадают в промежуточного хозяина при поедании им зрелых яиц паразита. Дальнейшее развитие цестод происходит с одной или двумя сменами промежуточных хозяев. Первым промежуточным хозяином цестод рыб обычно являются низшие ракообразные (чаще всего веслоногие), бокоплавы, мизиды, реже малощетинковые черви (у гвоздичников) и другие беспозвоночные. Яйца с онкосферой (у гвоздичников) или свободноплавающие корацидии (у ремнецов, ботриоцефалюса, триенофоруса и др.) заглатываются беспозвоночными животными, сквозь стенки их кишечника проникают в полость тела, превращаясь в процеркои-да — первую паразитическую стадию цестод. Процеркоид в полости тела промежуточного хозяина увеличивается в размерах; развиваются его внутренние системы (мышечная ткань, выделительная система). На заднем конце его образуется особый придаток — церкомер, в котором находятся зародышевые крючья. Дальнейшее развитие цестод различается в зависимости от того, с одним или с двумя промежуточными хозяевами протекает жизненный цикл гельминта. Если цикл развития происходит при участии одного промежуточного хозяина (гвоздичники, ботриоце-фалюс, протеоцефалюс), то уже у процеркоида формируются прикрепительные органы, характерные для взрослых червей, закладывается и развивается половая система. Попадая вместе с пищей в кишечник окончательного хозяина, процеркоид прикрепляется к стенкам кишечника и развивается во взрослого червя. Если цикл развития происходит при участии двух и более промежуточных хозяев, то процеркоид вместе с первым промежуточным хозяином (ракообразным) попадает в кишечник второго промежуточного хозяина, проникает в полость его тела (ремнецы), печень (триенофорус), мускулатуру (лентец широкий), где теряет церкомер и превращается в следующую личиночную стадию — плероцеркоид. Плероцеркоид растет. У ремнецов развиваются внутренние органы, заканчивается органогенез половой системы, у лентецов плероцеркоид слабо дифференцирован и внутренние органы его еще не заложены. Инвазионного состояния плероцеркоиды ремнецов достигают через 6—14 мес. Далее плероцеркоид вместе со вторым промежуточным хозяином попадает в кишечник окончательного хозяина, где достигает половой зрелости и продуцирует яйца. |