Дендрология в ВОПРОСАХ и ОТВЕТАХ 222222222. Дендрология в вопросах и ответах
 Скачать 4.08 Mb. Скачать 4.08 Mb.
|

|
Флористические области земного шара Основываясь на различиях растительности на поверхности суши земного шара, можно выделить шесть флористических областей (доминионы): неотропическая, палеотропическая, австралийская капская, антарктическая, голарктическая. Каждая флористическая область имеет свои не повторяющиеся вне ее семейства и роды растений, происхождение и распространение которых на протяжении геологической истории протекали в ее пределах. Расположение флористических областей во многом объясняет современное распределение ареалов растений, а также ареалов фитоценозов, в связи с климатическими и почвенными зонами. Чтобы уяснить особенности ареалов, рассмотрим краткую характеристику флористических областей суши (рис.). Голарктическая область занимает более половины суши земного шара. Для нее характерны многие семейства растении, свойственные умеренному, субарктическому и арктическому поясам: буковые, березовые, сосновые, ивовые, крестоцветные, зонтичные, осоковые, злаковые и др. Флора голарктической области происходит от неогеновой. Ядро ее составляет китайская флора, производная от тропической. Она является реликтовой и сохранилась до наших дней. Отличительной особенностью области является то, что наряду с элементами древней третичной флоры, из которой в дальнейшем развились субтропические, лиственные и хвойные леса, выделяется молодая флора — тундровая. Палеотропическая область характеризуется распространением различных родов пальм,из которых своеобразны: кокосовые, финиковые, борассовые, панданус («винтовая пальма»); гигантских трав из семейства банановых и бамбука, орхидей, влажных тропических гилей, баобабов и колючих кустарников, акаций саванн, суккулентов (алоэ, молочаи), растение вельвичия (в пустыне Калахари). В палеотропической области много эндемичных семейств и родов. К ним относятся вечнозеленые деревья тропических лесов: гигантские фикусы, масличные пальмы. Ядро этой флоры расположено в пределах Малайского архипелага. Неотропическая область. Южная граница проходит по 40 ю. ш. Отличается наибольшим богатством видов (в Бразилии насчитывается до 40 000). Оригинальность флоры выражается в обилии специфических семейств и родов растений таких, как кактусовые (центр распространения на Мексиканском нагорье и Бразильском плато), бромелиеивые (ананасные), канны (многие декоративные), виды пальм (например, кокосовые), орхидеи, папоротники (от миниатюрных мохоподобных до древовидных) и др. 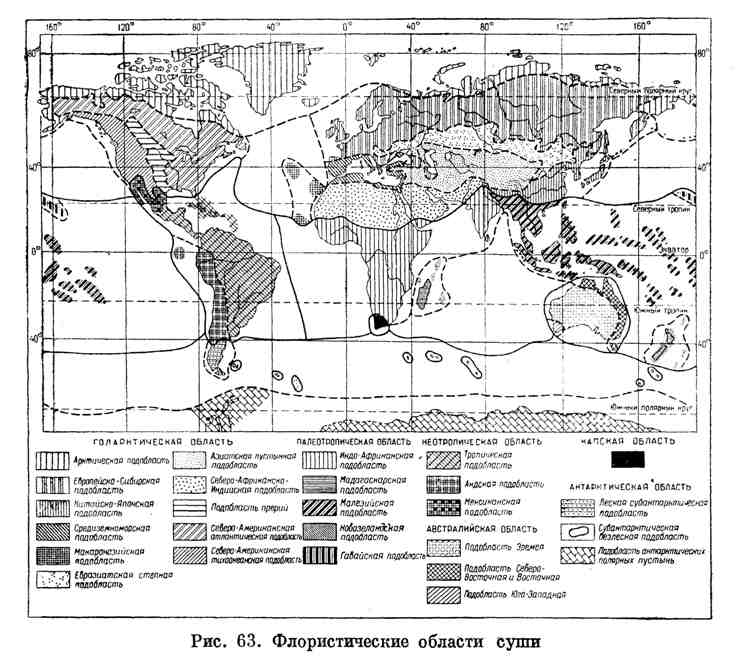 Рис. Флористические области суши Австралийская область характеризуется преобладанием эндемичных видов, которые приурочены к этому материку и имеют ограниченное распространение. По оригинальности и их обилию (до 75%) флористическая область не имеет равных. К числу эндемичных видов относятся эвкалипты, акации протейные, банксия,казуариновые (безлистные деревья) и др. Австралийская флора развивалась с мелового времени и сохранила свою специфичность вследствие географической изоляции. Капская область занимает юго-западную окраину Африки. Здесь распространено более 6000 видов, большая часть которых эндемична. Капская флора является реликтовой. В прошлые геологические эпохи она имела более широкое распространение (например, в субтропической Африке). Главными представителями капской флоры являются: вересковые(460 видов), протеиновые, ирисовые, орхидеи, пеларгонии, лилейные и др. Характерна бедность древесными видами. Капская флора имеет связи с австралийской. Антарктическая область невелика по площади и отличается от других бедностью флоры (на Огненной Земле насчитывается 615 видов, на материке 1 вид). Многие виды эндемичны: южный бук,из хвойных — араукарии, некоторые виды мхов. Ряд видов получает распространение в Капской и Австралийской области, что свидетельствует о связи антарктической области с другими материками. Встречаются также виды, свойственные северным субтропическим областям (водяника, примула, ясколка). Названные флористические области по своим границам нередко не совпадают или расходятся с современным распределением суши и моря. Так, палеотропическая область расположена между тропиками в восточном полушарии. Голарктическая — охватывает три материка: Северную Америку, северную Африку, Евразию. В этой области сохранились физико-географические условия, близкие к третичному времени, и богатейшая флора тропических лесов и саванн. Указанные флористические области позволяют выяснить многие особенности ареалов как отдельных видов, так и фитоценозов, что очень важно для их классификации. Растительные ареалы занимают большие площади. Контрастными являются космополитические и эндемичные ареалы. Космополитические ареалы занимают большую часть суши. Само понятие о растениях-космополитах в какой-то мере условно, так как каждое растение произрастает в определенных экологических условиях и оно не может «охватывать» всю сушу. К космополитам относятся многие культурные растения, сорные (осот, крапива, одуванчик), водные растения (тростник, камыш, водяные лилии), лесные виды (папоротник-орляк, плаун и др.). Эндемичные ареалы занимают незначительные территории, иногда площадью до нескольких квадратных километров. Растения присущи только одному району и в других местах не встречаются. Примером растений-эндемиков являются эвкалипты Австралии и Малайского архипелага, секвойи Калифорнии. Много эндемичных форм в горах (сосна Станкевича в Крыму, камчатская пихта), на островах Полинезии. Нередко эндемичные ареалы образуют реликтовыерастения, которые являются остатками древней угасшей флоры, имевшей развитие в предысторический период. Примеры таких реликтов — рододендрон и субальпийское высокотравье Кавказа. Сохранению реликтов способствует изолированность территории. ___________________________ 5. Понятие о виде. Вид как система популяций. Биоразнообразие и динамика экосистем: информационные технологии и моделирование. Отв. ред. В.К. Шумный и др. Новосибирск. Изд-во СО РАН. 2006. 648 с. ПОПУЛЯЦИЯ - 1. Совокупность особей одного вида, занимающих определенную территорию, свободно скрещивающихся друг с другом, имеющих общее происхождение, генетическую основу и в той или иной степени изолированная от других популяций данного вида. П. - элементарная эволюционная структура. Обладает рядом признаков, отсутствующих у отдельных ее членов. Контакты между особями внутри одной П. происходят чаще, чем между особями разных П. П. способна к развитию, в ней протекают простейшие эволюционные процессы. Локальная П. - географически отделенная в той или иной степени от др. группа особей одного вида. П. – Совокупность особей одного вида, длительно населяющих определенное пространство, внутри которого практически осуществляется та или иная степень панмиксии. П. отделена от соседних аналогичных совокупностей той или иной степенью давления различных форм изоляции и имеет сложную структуру по полу, возрасту, пространственным и близкородственным объединениям особей. П. – элементарная единица эволюционного процесса и форма существования вида. Каждая П. способна (теоретически – неограниченно долго) развиваться при подходящих условиях окружающей ее среды. Основными характеристиками П. являются: численность, плотность, рождаемость, смертность, прирост особей, темп роста и др. П. имеет определенную структуру: возрастную (соотношение особей разного возраста), сексуальную (соотношение полов) и пространственную (колонии, семьи, стаи). Так, возрастная структура П. является ее важной характеристикой, влияющей на рождаемость и смертность. Соотношение разных возрастных групп в П. определяет ее способность к размножению в данный момент, причем обычно в быстрорастущих П. значительную долю составляют молодые особи. П. реагируют на изменения внешней среды путем колебания численности, соотношения полов, возрастной структуры и т. д. Численность особей в П. колеблется в значительных пределах, однако она не может быть ниже некоторых пределов. Сокращение численности за эти пределы может привести к вымиранию П. Полагают, что при численности П. меньше нескольких сотен особей какие-либо случайные причины (пожар, наводнение, обильные снегопады, сильные морозы и т.п.) могут сократить численность настолько, что оставшиеся особи не смогут встретиться и оставить потомство. В соответствии с величиной популяционного ареала и численностью особей находится плотность особей в П. Соотношение половозрелых и неполовозрелых особей в П. различно и зависит от продолжительности жизни, времени наступления половой зрелости, интенсивности размножения. Генетический механизм определения пола обеспечивает расщепление потомства по полу в соотношении 1:1. В силу разной жизнеспособности мужских и женских организмов это первичное соотношение порой заметно отличается от вторичного (при родах – у млекопитающих) и тем более от третичного (характерного для взрослых особей). Например, в П. человека вторичное соотношение полов 100 девочек:106 мальчиков; к 16-18 годам это соотношение выравнивается и становится 1:1 (100 девочек:100 мальчиков), к 50 годам – 100 женщин:85 мужчин, а к 80 годам соотношение по полу становится 2:1 (100 женщин:50 мужчин). Размеры П. (пространственные и по числу особей) подвержены постоянным колебаниям. (Периодические колебания численности П. называются волнами жизни, или популяционными волнами.) Причины этих колебаний различны и в общей форме сводятся к влиянию биотических и абиотических факторов (влага, микроорганизмы, вызывающие заболевания, запас пищи, свет, температура, конкуренты, стихийные, бедствия и т.п.). Масштабы таких измерений могут быть весьма значительными. Так, например, на одном небольшом острове у юго-западного побережья Англии обитает в диком виде П. кроликов. В период максимальной численности (осенью благоприятного по кормовым условиям года) общее число кроликов достигало 10 тысяч особей, а однажды (после холодной и малокормной зимы) число их сократилось до 100 особей. С изменением числа особей в П. изменяется их плотность, т.е. число особей на единицу площади. (Определенная плотность П. поддерживается за счет конкуренции.) В отдельные годы резко увеличивается плотность П. насекомых-вредителей, кровососов, грызунов, сорных и др. растений. Верх, предел плотности П. определяется количеством самого дефицитного ресурса (воды или питательных элементов для растений, растительной пищи для травоядных животных, количеством жертв для хищников). Устойчивость П. поддерживается исторически сложившимися способами самовоспроизведения благодаря смене поколений и способности к саморегуляции путем изменения своей структуры. Например, в П. жука мучного хрущака при плотности П., превышающей оптимальную, внутривидовая конкуренция превращается в каннибализм – самцы поедают яйца, откладываемые самками, т.е. осуществляется изменение возрастной структуры П. и снижается ее плотность. У некоторых видов увеличение плотности П. вызывает резкое сокращение или вообще временную утрату способности давать потомство. У видов растений, не имеющих специальных приспособлений для распространения семян на большое расстояние, состояние перенаселенности (высокой плотности) возникает достаточно часто. В этих случаях, чтобы предотвратить очень резкую внутривидовую конкуренцию, уменьшается размер растений. Например, растения мари белой в зависимости от плотности П. могут отличаться по размерам в сотни раз. У этого вида развиваются практически все растения из всхожих семян. Однако меньшие по величине растения дадут и меньше семян. В результате, чем выше плотность П., тем меньше образуется семян, а в итоге плотность П. остается либо на прежнем уровне, либо снижается. В П. люцерны при плотности выше оптимальной растения не цветут и не плодоносят, но при этом происходит интенсивное накопление запасных питательных веществ в корневой шейке, которые используются для цветения и плодоношения, когда восстановится оптимальная плотность растений на единицу площади. Т. обр., регуляция численности и плотности особей в П. регулируется абиотическими и биотическими факторами, в т. ч. конкуренцией, а также внутрипопуляционными механизмами. Судьба вида в биоценозе. Виды, составляющие каждый конкретный биоценоз, различаются своими требованиями к окружающей среде. Поэтому биологические особенности вида лежат в основе его экологической индивидуальности в структуре биоценоза. Своеобразие видов проявляется в различных требованиях к вещественным и энергетическим условиям среды, необходимым для роста и развития, в темпах размножения как основной функции, обеспечивающей положение вида в сообществе. Попытки объяснить судьбу вида в биоценозе привели к формированию концепций стратегий вида и концепции экологической ниши. В начале XX в. Л.Г. Раменский сформулировал концепцию экологической индивидуальности вида, основанную на идее конкурентных отношений за вещественные и энергетические ресурсы в каждом конкретном месте земной поверхности. Ему удалось выделить три основных типа стратегии вида в биоценозе. 1. Виды, обладающие высокой конкурентоспособностью и энергией жизнедеятельности, полнотой использования ресурсов среды, названные им виолентами (более образно — «львами»). Это абсолютные конкуренты, способные вступать в борьбу за жизненный успех со всеми другими видами. Число их, видимо, невелико. 2. Виды, обладающие низкой конкурентоспособностью, но зато быстро осваивающие новые территории. Они получили название эксплерентов (или «шакалов»). Это пионерные виды. Они быстро заселяют свободные территории, но так же быстро и уходят с них. Число их также невелико. 3. Виды, обладающие большой выносливостью в различных условиях, получили название патиентов («верблюдов»). Это основная масса видов. Различные группы патиентов могут обладать чертами как виолентов, так и эксплерентов. Т. А. Работнов, разрабатывая данную концепцию, пришел к выводу, что виолентность, патиентность, эксплерентность присущи не только виду в целом, но проявляются и на уровне отдельных популяций и особей. Э. Пианка предложил другой подход к выделению стратегий вида, основанный на учете доли энергии, используемой на размножение, от общих затрат на жизнедеятельность. В рамках этой концептуальной основы появились представленияоk- и r-стратегиях. Общие характеристики видов, обладающих такими стратегиями, приведены в табл. 5.5. Дальнейшая разработка этой концепции привела к выделению различных промежуточных вариантов с различным сочетанием свойств обеих стратегий. В общем плане r-стратегия характеризуется большим энергетическим вкладом в производство потомства, а k-стратегия — затратами на поддержание жизненной активности, т. е. на рост и поддержание во взрослом состоянии. Другая концепция основана на понятии экологической ниши. Термин «ниша» впервые применил для обозначения самой малой единицы распространения вида Р. Джонсон в 1910 г. Через 17 лет, в 1927 г., Ч. Элтон отметил функциональный аспект этого термина и определил его как место вида в структуре биоценоза. Идея экспериментально исследована Г. Ф. Гаузе, Т. Парком и др., что позволило сформулировать эмпирическое обобщение: «Полные конкуренты не могут существовать бесконечно». Из обобщения следует, что если два вида существуют вместе, то между ними должно быть какое-то биологическое различие, обусловливающее различные экологические ниши обоих видов. Таблица Общая характеристика видов, обладающих разными стратегиями
Д. Хатчисон (1958) считал, что нишу следует определить в рамках всего диапазона физических, химических, биотических переменных среды. В идеале каждую такую переменную можно рассматривать как некий градиент, на котором у каждого вида имеется свой диапазон активности. Ю. Одум (1971) настаивает на придании нише функционального аспекта («профессии» вида в биоценозе), оставив остальные аспекты в рамках понятия «местообитание». В общем плане экологическая ниша — это процесс дифференцировки пространственно-временных отношений между видами, составляющими конкретный биоценоз. Теоретически, при исключении всяких давлений извне, каждый вид способен освоить всю совокупность условий в конкретном месте. Это его потенциальная, или фундаментальная, ниша. Наличие других видов не позволяет ее реализовать в полной мере. Поэтому реальная экологическая ниша всегда будет частью фундаментальной. В ходе процесса дифференцировки пространства, занятого биоценозом, экологические ниши видов могут проявляться как независимые, с разной степенью перекрытия, и ниша одного вида может находиться внутри ниши другого. Концепции экологической индивидуальности вида и экологической ниши, рассматривая разные стороны судьбы вида в биоценозе, дополняют друг друга. Остается неразработанным вопрос о роли стратегий видов в формировании структуры биоценоза: как виды с разной стратегией формируют ниши, чем отличаются экологические ниши видов разных стратегий, за счет особей каких стратегий решаются те или иные вопросы взаимоотношений с окружающим миром. Устойчивость биоценоза является характеристикой, наиболее интересной для оценки его связей с окружающей средой, т. к. она выступает как высший интегральный показатель всех процессов, протекающих внутри биоценоза. До 70-х гг. XX в. считалось, что устойчивость сообщества определяется видовым разнообразием: чем больше видов, тем устойчивей биоценоз к различным внешним воздействиям. Теоретическое осмысление этой идеи, разработанные разнообразные математические модели, экспериментальные исследования и наблюдения в природе привели к формированию трех основных взглядов на данную проблему: • устойчивость биоценоза повышается с увеличением видового разнообразия; • устойчивость биоценоза снижается с увеличением видового разнообразия; • устойчивость всех биоценозов Земли примерно одинакова, т. е. является некоторой константой. Каждая из этих точек зрения имеет своих сторонников и противников и, что особенно важно отметить, определенную систему доказательств: от теоретических посылок, математических моделей до наблюдаемых явлений в природе. М. Бигон, проведя достаточно большой анализ данной проблемы, заканчивает его следующими словами: ...Ясно одно: такой вещи, как устойчивость сообщества, вообще попросту не существует. Этот параметр зависит от исследуемого компонента системы и характера нарушения. Рассмотрение биоценоза как биотической компоненты экосистемы позволяет несколько прояснить этот вопрос. Поскольку экосистема есть мыслительная конструкция, выделяемая каждый раз для решения конкретной задачи, то и характеристика ее устойчивости определяется также в рамках задачи. |