биохимия лекции. Энзимология как учение о ферментах. Простые и сложные ферменты
 Скачать 3.9 Mb. Скачать 3.9 Mb.
|

|
Раздел 3.1 Структура коферментов. Участие витаминов в построении коферментов. Коферменты относятся к сложным органическим веществам, их молекулы значительно меньше по размеру, чем молекулы ферментов. Коферменты могут проникать через биологические мембраны, нагревание обычно не вызывает изменения их структуры. Функцией кофермента является участие в катализируемой реакции, причём количество фермента и его химическое строение внешне остаются неизменными. В действительности кофермент является одним из субстратов ферментативной реакции, т.е. выступает как косубстрат. В ходе реакции кофермент претерпевает химические превращения, в точности противоположные тем, которые происходят в субстрате. Например, в окислительно-восстановительных реакциях молекула субстрата окисляется, а молекула кофермента восстанавливается. При последующих сопряжённых реакциях изменения в коферменте протекают в обратном направлении и он воспроизводится в первоначальной форме. Таким образом, коферменты могут быть охарактеризованы как переносчики определённых атомов, электронов или химических групп на соответствующий акцептор. Строение апофермента определяет специфичность этой реакции, а строение кофермента – её тип. Классификация коферментов. По происхождению и химическому строению коферменты можно подразделить на витаминсодержащие (витаминные) и невитаминные. К первой группе относятся производные водорастворимых витаминов группы В. В их состав могут входить также адениловые нуклеотиды. Особенности структуры и функции витаминных коферментов приведены в таблице. К невитаминным коферментам относятся в первую очередь пептидные и нуклеотидные коферменты. Глутатион – кофермент пептидной природы. По химическому строению это трипептид γ–глутамил-цистеинил-глицин. Его реакционная способность определяется SH-группой цистеина, которая легко вступает в окислительно-восстановительные реакции. Поэтому глутатион может находиться в восстановленной (Г-SH) и окисленной (Г-S-S-Г) форме. В клетке глутатион присутствует преимущественно в восстановленной форме. Его основная функция состоит в том, что глутатион защищает SH-группы ферментов от окисления и образует тиолы из дисульфидов. 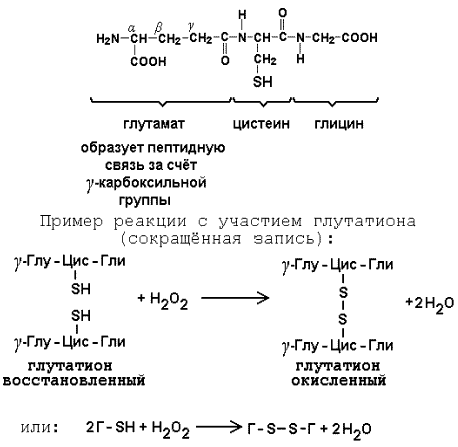 Аденозинтрифосфат (АТФ) - кофермент нуклеотидной природы. В его состав входит пуриновое основание аденин, углевод рибоза и три остатка фосфорной кислоты. Это соединение содержит богатые энергией (макроэргические) фосфатные связи и может принимать участие в реакциях синтеза сложных веществ, а также служить донором фосфатной группы. 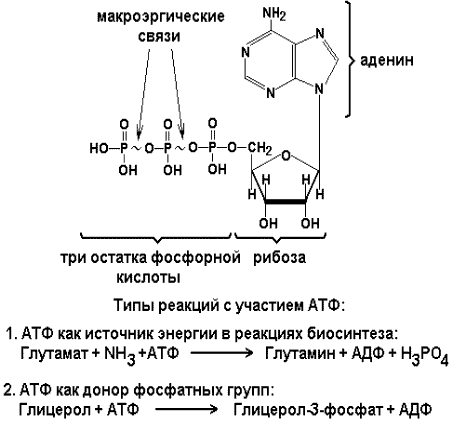 Небелковая часть сложного фермента может быть представлена также ионами металлов. Ферменты, содержащие в своём составе ионы металлов, называются металлоферментами (например, α-амилаза включает в свой состав ионы Са2+, которые участвуют в стабилизации пространственной структуры фермента). Удаление этих ионов приводит к потере активности фермента. В других случаях ионы металлов могут служить активаторами ферментов - реакция может происходить и в отсутствие этих ионов. Многие коферменты и простетические группы ферментов являются производными витаминов — органических веществ, которые не синтезируются в организме человека и должны поступать в составе пищевых продуктов. Они называются витаминными коферментами. В молекуле такого кофермента активным компонентом, соединяющимся с переносимой группой, служит именно витамин. Остальная часть молекулы кофермента обеспечивает специфическое связывание с апоферментом в строго определённой ориентации. Заболевания, возникающие у людей при недостатке витаминов в пище, являются следствием нарушений обмена веществ, в результате снижения концентрации коферментов специфических ферментативных реакций. Важнейшими из витаминных коферментов являются: Тиаминдифосфат (ТДФ) является производным витамина В1; участвует в реакциях окислительного декарбоксилирования пировиноградной и α-кетоглутаровой кислот. Его формула приведена на рисунке. 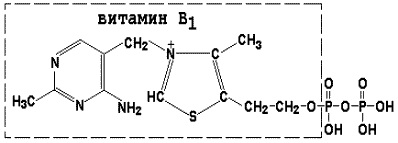 Пиридоксальфосфат является производным витамина В6; он принимает участие в реакциях трансаминирования аминокислот. Его формула приведена на рисунке. 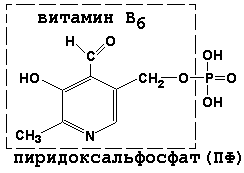 Коферменты НАД+ (никотинамидадениндинуклеотид) и НАДФ+ (никотинамидадениндинуклеотидфосфат) содержат в своём составе витамин РР и принимают участие в окислительно-восстановительных реакциях. Восстановленные формы этого кофермента обозначается НАДН и НАДФН соответственно. Формула кофермента НАД+ представлена на рисунке; структура НАДФ+ отличается наличием дополнительной фосфатной группы во втором положении рибозы аденилового нуклеотида. 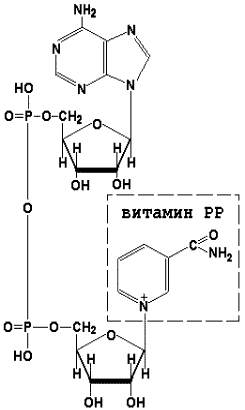 Образование восстановленных форм никотинамидных коферментов представлено на схеме: 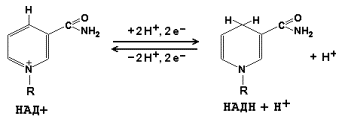 Коферменты ФАД (флавинадениндинуклеотид) и ФМН (флавинмононуклеотид) содержат в своём составе витамин В2 и принимают участие в окислительно-восстановительных реакциях. Восстановленные формы этого кофермента обозначается ФАДН2 и ФМНН2 соответственно.  Коэнзим А (КоА-SH) является производным витамина В3 (пантотеновой кислоты) и участвует в реакциях переноса остатков жирных кислот (реакциях ацилирования). 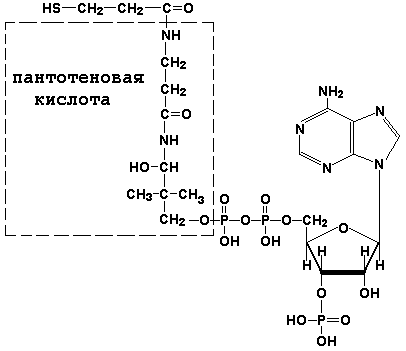 5,6,7,8-Тетрагидрофолиевая кислота (ТГФК) является производным витамина Вс (фолиевой кислоты) и участвует в реакциях переноса одноуглеродных групп: метильной (СН3-), метиленовой (-СН2-), метенильной (-СН=), формильной (-СОН) и некоторых других. 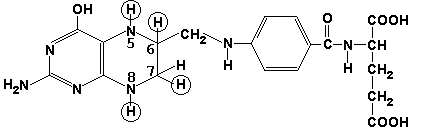 Метилкобаламин (содержит витамин В12) и также принимает участие в переносе метильной группы.  Биотин (витамин Н) участвует в активации СО2 и переносе карбоксильных групп (реакциях карбоксилирования). 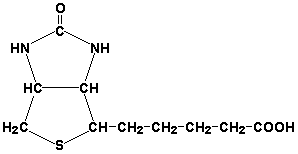 В настоящее время известно более двух тысяч химических реакций, катализируемых ферментами, и число это непрерывно возрастает. Чтобы ориентироваться в таком множестве превращений. возникла настоятельная необходимость в систематизированной классификации и номенклатуре, при помощи которой любой фермент можно было бы точно идентифицировать. Номенклатура, которой пользовались до середины XX века, была весьма далека от совершенства. Исследователи, открывая новый фермент, давали ему название по своему усмотрению, что неизбежно вело к путанице и всевозможным противоречиям. Некоторые названия оказались ошибочными, другие ничего не говорили о природе катализируемой реакции. Учёные разных школ часто употребляли разные названия для одного и того же фермента или, наоборот, одно и то же название для нескольких разных ферментов. Было решено разработать рациональную международную классификацию и номенклатуру ферментов, которой могли бы пользоваться биохимики всех стран. С этой целью при Международном союзе биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology, IUВMB) была создана Комиссия по ферментам, предложившая в 1964 году основные принципы такой классификации и номенклатуры. Она постоянно совершенствуется и дополняется, в настоящее время действует уже шестая редакция этой номенклатуры (1992 год), к которой ежегодно выходят дополнения. Рекомендации Комиссии по ферментам можно найти в Интернете (рисунок 3.1) по адресу: http://www.chem.qmul.ac.uk/iubmb/enzyme/index.html . Рисунок 3.1. Стартовая страница сайта Комиссии по ферментам Международного союза биохимии и молекулярной биологии. В основу классификации положен важнейший признак, по которому один фермент отличается от другого — это катализируемая им реакция. Число типов химических реакций сравнительно невелико, что позволило разделить все известные в настоящее время ферменты на 6 важнейших классов, в зависимости от типа катализируемой реакции. Такими классами являются: оксидоредуктазы (окислительно-восстановительные реакции); трансферазы (перенос функциональных групп); гидролазы (реакции расщепления с участием воды); лиазы (разрыв связей без участия воды); изомеразы (изомерные превращения); лигазы (синтез с затратой молекул АТФ). Ферменты каждого класса делят на подклассы, руководствуясь строением субстратов. В подклассы объединяют ферменты, действующие на сходно построенные субстраты. Подклассы разбивают на подподклассы, в которых ещё строже уточняют структуру химических групп, отличающих субстраты друг от друга. Внутри подподклассов перечисляют индивидуальные ферменты. Все подразделения классификации имеют свои номера. Таким образом, любой фермент получает свой уникальный кодовый номер, состоящий из четырёх чисел, разделённых точками. Первое число обозначает класс, второе - подкласс, третье - подподкласс, четвёртое - номер фермента в пределах подподкласса. Например, фермент α-амилаза, расщепляющая крахмал, обозначается как 3.2.1.1, где: 3 — тип реакции (гидролиз); 2 — тип связи в субстрате (гликозидная); 1 — разновидность связи (О-гликозидная); 1 — номер фермента в подподклассе (рисунок 3.2). Рисунок 3.2. Страница сайта Комиссии по ферментам, содержащая информацию об α-амилазе. Вышеописанный десятичный способ нумерации имеет одно важное преимущество: он позволяет обойти главное неудобство сквозной нумерации ферментов, а именно: необходимость при включении в список вновь открытого фермента изменять номера всех последующих. Новый фермент может быть помещён в конце соответствующего подподкласса без нарушения всей остальной нумерации. Точно так же при выделении новых классов, подклассов и подподклассов их можно добавлять без нарушения порядка нумерации ранее установленных подразделений. Если после получения новой информации возникает необходимость изменить номера некоторым ферментам, прежние номера не присваивают новым ферментам, чтобы избежать недоразумений. Говоря о классификации ферментов, следует также отметить, что ферменты классифицируются не как индивидуальные вещества, а как катализаторы определённых химических превращений. Ферменты, выделенные из разных биологических источников и катализирующие идентичные реакции, могут существенно отличаться по своей первичной структуре. Тем не менее в классификационном списке все они фигурируют под одним и тем же кодовым номером. Знание кодового номера фермента позволяет: устранить неоднозначности, если разные исследователи используют одно и то же название для различных ферментов; сделать поиск информации в литературных базах данных более эффективным; получить в других базах данных дополнительную информацию о последовательности аминокислот, пространственной структуре фермента, генах, кодирующих ферментные белки. Система классификации, разработанная Комиссией по ферментам, включает также и вновь созданную номенклатуру ферментов, которая строится по специальным принципам. Согласно рекомендациям IUBMB, ферменты получают два рода названий: систематическое и рабочее (рекомендуемое). Систематическое название составляется из двух частей. Первая часть содержит название субстрата или субстратов, часто — наименование кофермента, вторая часть указывает на природу катализируемой реакции и включает название класса, к которому относится данный фермент. При необходимости приводится дополнительная информация о реакции в скобках после второй части названия. Систематическое название присваивается только тем ферментам, каталитическое действие которых полностью изучено. Например, систематическое название α-амилазы — 1,4-α-D-глюкан-глюканогидролаза. Конечно, такое название очень неудобно для запоминания и произнесения. Поэтому наряду с систематическими Комиссия по ферментам IUBMB даёт рекомендует использовать рабочие (упрощённые) названия ферментов. Рабочее название фермента должно быть достаточно коротким для употребления. В качестве рабочего названия в ряде случаев может быть использовано тривиальное название, если оно не является ошибочным или двусмысленным. В других случаях оно строится на тех же общих принципах, что и систематическое название, но с минимальной детализацией. Конкретные примеры систематических и рабочих названий ферментов приводятся в следующем разделе данной темы курса. В научных публикациях при первом упоминании о ферменте принято указывать его систематическое название и кодовый номер, а в дальнейшем пользоваться его рабочим названием. Основные правила построения систематических и рабочих названий разных классов ферментов: Оксидоредуктазы Систематическое название ферментов этого класса строится по схеме донор: акцептор - оксидоредуктаза. Согласно тривиальной номенклатуре, оксидоредуктазы, отщепляющие атомы водорода или электроны и переносящие их на любой акцептор, кроме кислорода, называются дегидрогеназами. Оксидоредуктазы, использующие кислород в качестве акцептора атомов водорода или электронов, называются оксидазами. Некоторые ферменты, которым свойственно преимущественно восстанавливающее действие, носят название редуктаз. Все перечисленные наименования могут быть использованы для построения рабочего названия оксидоредуктаз. Трансферазы Систематическое название ферментов, ускоряющих такие реакции, составляют по форме донор:акцептор (транспортируемая группа) трансфераза. В рабочем названии обычно указывается только один специфический субстрат или продукт наряду с названием транспортируемой группировки. Гидролазы Систематическое название составляется по форме субстрат-гидролаза. У гидролаз, специфически отщепляющих определённую группу, эта группа может быть указана в виде префикса. Рабочее название чаще всего составляется из названия гидролизуемого субстрата с добавлением окончания -аэа. Следует, однако, отметить, что вследствие достаточно сложного и зачастую до конца не выявленного характера специфичности многих гидролаз не всегда удаётся дать им систематическое название. В этих случаях рекомендовано использовать эмпирические названия, присвоенные им при первом описании. Так, не имеют систематического названия такие ферменты, как пепсин, папаин, тромбин. Лиазы Систематическое название ферментов строится по схеме: субстрат-отщепляемая группа-лиаза. Чтобы уточнить, какая группа отщепляется, используются префиксы "карбокси-", "аммиак", "гидро-" и т.д. В качестве рабочих названий ферментов сохраняются тривиальные названия типа "декарбоксилаза", "альдолаза", "дегидратаза", "десульфгидраза". Лиазы делятся на подклассы в зависимости от характера разрываемых связей. Изомеразы Систематическое название ферментов включает название субстрата и слово изомераза, которому предшествует указание типа реакции изомеризации. Рабочие названия подобны (с некоторыми упрощениями) систематическим названиям. Лигазы Систематическое название образуется из названий соединяемых субстратов в сочетании со словом лигаза. В скобках указывается продукт, образующийся в результате гидролиза нуклеозидтрифосфата. Рабочее название ферментов этого класса составляется, как правило, из названия продукта реакции в сочетании со словом синтетаза. Знакомясь в последующем с различными ферментативными реакциями, всегда анализируйте сущность изменений, происходящих в субстратах, и пытайтесь определить по крайней мере класс фермента, катализирующего реакцию. Анализируйте также названия ферментов и соотносите их с процессами, происходящими в реакциях. Это облегчит запоминание названий ферментов и катализируемых ими превращение и позволит больше времени уделить уяснению биологической роли изучаемых процессов. Раздел 2.3 Характеристика классов ферментов, формирование подклассов и подподклассов. 1. ОКСИДОРЕДУКТАЗЫ К классу оксидоредуктаз относят ферменты, катализирующие окислительно-восстановительные реакции. Общая схема их может быть представлена следующим образом: , где AH2 —донор водорода, B — акцептор водорода. В живых организмах окисление осуществляется преимущественно путём отщепления атомов водорода или электронов от субстратов-доноров. Акцепторами атомов водорода или электронов могут быть различные вещества - коферменты (НАД, НАДФ, ФАД, ФМН, глутатион, липоевая кислота, убихинон), цитохромы. железосерные белки и кислород. Подклассы оксидоредуктаз формируются в зависимости от природы функциональной группы донора водорода (электронов). Всего выделяют 19 подклассов. Основными из них являются следующие: Оксидоредуктазы, действующие на СН-ОН-группу доноров. Ферменты, относящиеся к этому подклассу, окисляют спиртовые группы до альдегидных или кетонных групп. В качестве примера можно привести фермент алкогольдегидрогеназу (алкоголь:НАД-оксидоредуктаза; КФ 1.1.1.1). участвующую в метаболизме этанола в тканях: 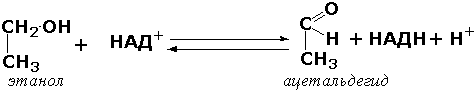 Кроме окисления спиртов, ферменты этого подкласса участвуют в дегидрировании оксикислот (молочной, яблочной, изолимонной), моносахаридов и других соединений, содержащих гидроксильные группы. Оксидоредуктазы, действующие на альдегидную или кетонную группу доноров. Эти ферменты окисляют альдегиды и кетоны до карбоновых кислот. К примеру, представитель данного подкласса - глицеральдегид-3-фосфатдегидрогеназа (D-глицеральдегид-3-фосфат:НАД-оксидоредуктаза (фосфорилирующая), КФ 1.2.1.12) - катализирует одну из промежуточных реакций распада глюкозы: 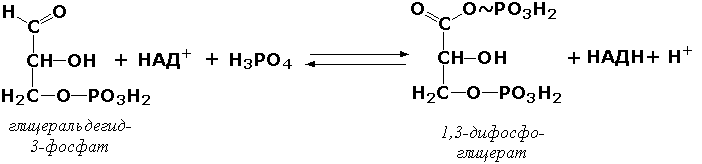 Важно отметить, что продукт этой реакции содержит богатую энергией фосфатную связь в 1-ом положении. Остаток фосфорной кислоты, образующий эту связь, может быть перенесён от 1,3-дифосфоглицерата на АДФ с образованием АТФ (см. далее). Оксидоредуктазы, действующие на СН-СН-группу доноров. В результате катализируемых ими реакций СН-СН-группы превращаются в С=С-группы. то есть происходит образование ненасыщенных соединений из насыщенных. Например, фермент цикла трикарбоновых кислот сукцинатдегидрогеназа (сукцинат:акцептор - оксидоредуктаза, КФ 1.3.99.1) ускоряет окисление янтарной кислоты с образованием ненасыщенной фумаровой кислоты:  Оксидоредуктазы, действующие на CH-NH2-группу доноров. Эти ферменты катализируют окислительное дезаминирование аминокислот и биогенных аминов. Амины при этом превращаются в альдегиды или кетоны, аминокислоты - в кетокислоты и выделяется аммиак. Так, глутаматдегидрогеназа (L-глутамат:НАД(Ф) - оксидоредуктаза (дезаминирующая), КФ 1.4.1.3) принимает участие в следующем превращении глутамата: 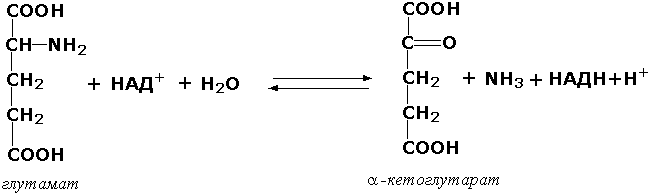 Оксидоредуктазы, действующие на серосодержащие группы доноров, катализируют окисление тиоловых (сульфгидрильных) групп до дисульфидных, а сульфитов - до сульфатов. Примером фермента является дигидролипоилдегидрогеназа (КФ 1.8.1.4), катализирующая одну из промежуточных реакций окислительного декарбоксилирования пирувата:  Оксидоредуктазы, действующие на пероксид водорода в качестве акцептора, сравнительно немногочисленны и объединены в отдельный подкласс, известный также под тривиальным названием пероксидазы. Примером фермента является глутатионпероксидаза (глутатион:Н2О2 - оксидоредуктаза. КФ 1.11.1.9), участвующая в инактивации пероксида водорода в эритроцитах, печени и некоторых других тканях: 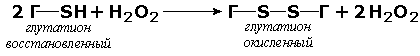 Оксидоредуктазы, действующие на пару доноров с включением молекулярного кислорода, или монооксигеназы - ферменты, катализирующие окисление органических соединений молекулярным кислородом, приводящее к включению одного из атомов кислорода в молекулы этих соединений. При этом второй атом кислорода включается в молекулу воды. Так реакция превращения фенилаланина в тирозин катализируется фенилаланин-4-монооксигеназой (КФ 1.14.16.1): 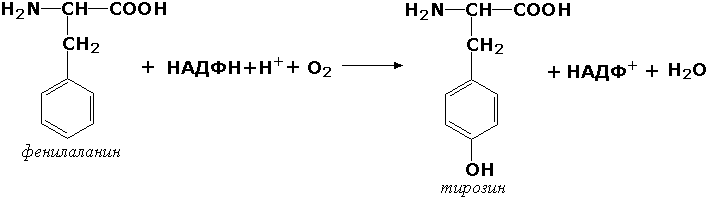 У некоторых людей генетический дефект этого фермента служит причиной заболевания, которое носит название фенилкетонурии. К монооксигеназам относится также фермент, известный под названием цитохром Р450 (КФ 1.14.14.1) Он содержится, главным образом, в клетках печени и осуществляет гидроксилирование чуждых организму липофильных соединений, образующихся в качестве побочных продуктов реакций или попадающих в организм извне. Например, индол, образующийся из триптофана в результате деятельности микроорганизмов кишечника, подвергается в печени гидроксилированию по следующей схеме: 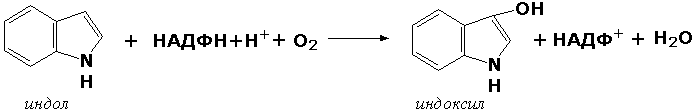 Появление гидроксильной группы повышает гидрофильность веществ и облегчает их последующий вывод из организма. Кроме того, цитохром Р450 принимает участие в отдельных этапах превращения холестерина и стероидных гормонов. Наличие у живых организмов высокоэффективной системы цитохрома Р450 приводит в ряде случаев к нежелательным практическим следствиям: сокращает время пребывания в организме человека лекарственных препаратов и тем самым снижает их терапевтический эффект. Оксидоредуктазы, действующие на один донор с включением молекулярного кислорода, или диоксигеназы, катализируют превращения, в ходе которых оба атома молекулы О2 включаются в состав окисляемого субстрата. Например, в процессе катаболизма фенилаланина и тирозина происходит образование из гомогентизиновой кислоты малеилацетоацетата, в состав которого включаются оба атома кислорода:  Фермент, катализирующий эту реакцию, называется гомогентизат-1,2-диоксигеназой (КФ 1.13.11.5). В ряде случаев встречается врождённый дефицит этого фермента, что приводит к развитию заболевания, называемого алкаптонурией. II. ТРАНСФЕРАЗЫ Трансферазы - класс ферментов, катализирующих перенос функциональных групп от одного соединения к другому. В общем виде эти превращения можно записать: где Х - переносимая функциональная группа. AX - донор группировки, В - акцептор. Подразделение на подклассы зависит от природы переносимых группировок. Трансферазы, переносящие одноуглеродные фрагменты. К этому подклассу относятся ферменты, ускоряющие перенос метильных (—CH3), метиленовых (—СН2—), метенильных (—СН=), формильных и родственных им групп. Так, при участии гуанидинацетат-метилтрансферазы (S-аденозилметионингуанидинацетат-метилтрансфераза, КФ 2.1.1.2) происходит синтез биологически активного вещества креатина: 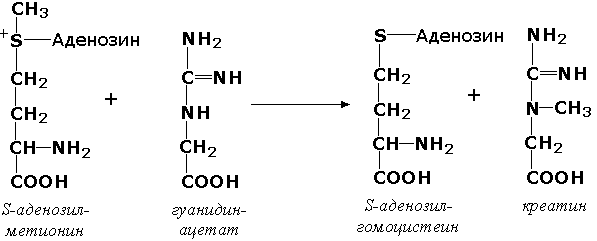 Трансферазы, переносящие остатки карбоновых кислот (ацилтрансферазы). Они катализируют разнообразные химические процессы связанные с переносом остатков различных кислот (уксусной, пальмитиновой и др.) преимущественно от тиоэфиров коэнзима А на различные акцепторы. Примером реакции трансацетилирования может быть образование медиатора ацетилхолина при участии холин-ацетилтрансферазы (ацетил-КоА:холин-О-ацетилтрансфераза, КФ 2.3.1.6):  Трансферазы, переносящие гликозильные остатки (гликозилтрансфсразы), катализируют транспорт гликозильных остатков из молекул фосфорных эфиров к молекулам моносахаридов, полисахаридов и других веществ. Эти ферменты, в частности, играют основную роль в синтезе гликогена и крахмала, а также в первой фазе их деструкции. Ещё один фермент этого подкласса - УДФ-глюкуронилтрансфераза (УДФ-глюкуронат-глюкуронил-трансфераза (неспецифичная к акцептору), КФ 2.4.1.17) - участвует в процессах обезвреживания эндогенных и чужеродных токсических веществ в печени: 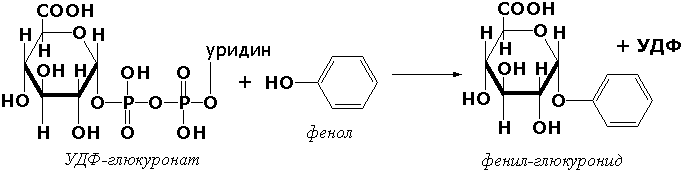 Трансферазы, переносящие азотистые группы. В этот подкласс входят аминотрансферазы, ускоряющие перенос α-аминогруппы аминокислот к α-углеродному атому кетокислот. Наиболее важным из этих ферментов является аланинаминотрансфераза (L-аланин:2-оксоглутарат-аминотрансфераза, КФ 2.6.1.2). катализирующая реакцию: 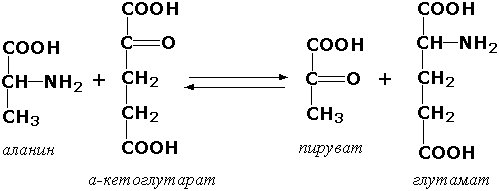 Трансферазы, переносящие фосфатные группы (фосфотрансферазы). Эта группа ферментов катализирует биохимические процессы, связанные с транспортом остатков фосфорной кислоты на различные субстраты. Указанные процессы имеют важное значение для жизнедеятельности организма, так как обеспечивают превращение ряда веществ в органические фосфоэфиры, обладающие высокой химической активностью и легко вступающие в последующие реакции. Фосфотрансферазы, использующие в качестве донора фосфата АТФ, принято называть киназами. Широко распространённым ферментом является гексокиназа (ATФ:D-гексоза-6-фосфотрансфераза. КФ 2.7.1.1.), ускоряющая перенос фосфатной группы с АТФ на моносахариды:  В некоторых случаях возможен и обратный перенос фосфатной группы с субстрата на АДФ с образованием АТФ. Так, фермент фосфоглицераткиназа (АТФ:D-3-фосфоглицерат-1-фосфотрансфераза, КФ 2.7.2.3) осуществляет превращение упомянутого ранее (см. "Оксидоредуктазы") 1.3-дифосфоглицерата:  Подобные реакции фосфорилирования АДФ с образованием АТФ, сопряжённые с превращением субстрата (а не с переносом электронов в дыхательной цепи), получили название реакций субстратного фосфорилирования. Роль этих реакций в клетке значительно возрастает при недостатке кислорода в тканях. III. ГИДРОЛАЗЫ Гидролазы - класс ферментов, катализирующих реакции расщепления органических соединений при участии воды (реакции гидролиза). Эти реакции протекают по следующей схеме: , где А-В - сложное соединение, А-Н и В-ОН - продукты его гидролиза. Реакции этого типа активно протекают в организме; они идут с выделением энергии и, как правило, необратимы. Подклассы гидролаз формируются в зависимости от типа гидролизуемой связи. Наиболее важными являются следующие подклассы: Гидролазы, действующие на сложные эфиры (или эстеразы) гидролизуют сложные эфиры карбоновой, фосфорной, серной и других кислот. Широко распространённым ферментом этого подкласса является триацилглицероллипаза (гидролаза эфиров глицерола, КФ 3.1.1.3). ускоряющая гидролиз ацилглицеролов: 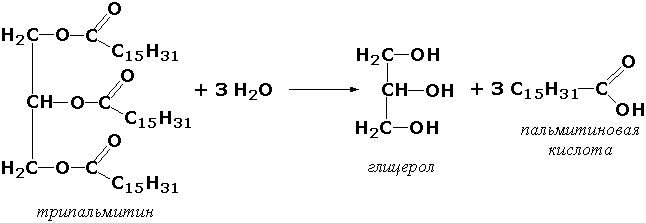 Другие представители эстераз расщепляют сложноэфирные связи в ацетилхолине (ацетилхолинэстераза), фосфолипидах (фосфолипазы), нуклеиновых кислотах (нуклеазы), фосфоорганических эфирах (фосфатазы). Гидролазы, действующие на гликозидные связи (гликозидазы) ускоряют реакции гидролиза олиго- и полисахаридов, а также других соединений, содержащих моносахаридные остатки (например, нуклеозидов). Характерным представителем является сахараза (β-D-фруктофуранозид-фруктогидролаза, КФ 3.2.1.26). катализирующая расщепление сахарозы: 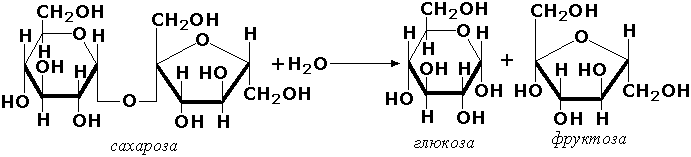 Гидролазы, действующие на пептидные связи (пептидазы), катализируют реакции гидролиза пептидных связей в белках и пептидах. К этой группе относятся пепсин, трипсин, химотрипсин, катепсин и другие протеолитические ферменты. Гидролиз пептидных связей происходит по следующей схеме:  Гидролазы, действующие на C-N-связи, отличающиеся от пептидных, - ферменты, ускоряющие гидролиз амидов органических кислот. Представитель этого подкласса - глутаминаза (L-глутамил-амидогидролаза, КФ 3.5.1.2) - участвует в поддержании кислотно-основного состояния организма, катализируя в почках гидролиз глутамина:  IV. ЛИАЗЫ Лиазы - класс ферментов, катализирующих негидролитические реакции расщепления субстратов с образованием двойных связей или, наоборот, присоединения по месту разрыва двойной связи. Общая схема этих реакций: , где А—В - субстрат, А и В - продукты реакции. В результате таких реакций часто выделяются простые вещества, например, СО2, NH3, H2О. Углерод-углерод-лиазы катализируют разрыв связи между двумя атомами углерода. Среди них наибольшее значение имеют карбокси-лиазы (декарбоксилазы), под влиянием которых осуществляется декарбоксилирование a-кето- и аминокислот, лиазы кетокислот , к которым относится цитратсинтаза, альдегид-лиазы (альдолазы). К последним относится фруктозодифосфатальдолаза (фруктозо-1,6-дифосфат-D-глицеральдегид-З-фосфат-лиаза, КФ 4.1.2.13), катализирующая реакцию: 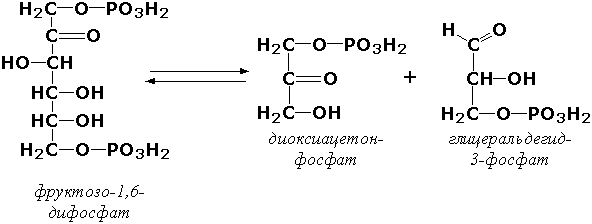 Углерод-кислород-лиазы катализируют разрыв связи между атомами углерода и кислорода. В этот подкласс входят прежде всего гидро-лиазы, участвующие в реакциях дегидратации и гидратации. Примером может служить сериндегидратаза (L-серин-гидро-лиаза (дезаминирующая), КФ 4.2.1.3), осуществляющая превращение: 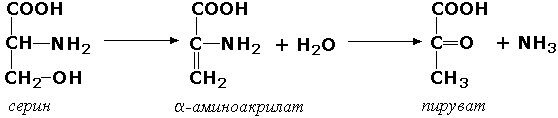 Иногда за основу рабочего названия может быть принята обратная реакция с применением термина "гидратаза". Так, для фермента цикла трикарбоновых кислот L-малат-гидро-лиазы (КФ 4.2.1.2) рекомендовано название "фумаратгидратаза":  Углерод-азот-лиазы участвуют в отщеплении азотсодержащих групп. Представителем этого подкласса является гистидин-аммиак-лиаза (L-гистидин-аммиак-лиаза, КФ 4.3.1.3), участвующая в дезаминировании гистидина:  Углерод-сера-лиазы катализируют отщепление сульфгидрильных групп. К этому подклассу относятся десульфгидразы серу содержащих аминокислот, например, цистеиндесульфгидраза (L-цистеин-сероводород-лиаза (дезаминирующая), КФ 4,4.1.1). V. ИЗОМЕРАЗЫ Изомеразы - класс ферментов, ускоряющих процессы внутримолекулярных превращений с образованием изомеров. Схематически реакции такого типа могут быть представлены следующим образом: , где А и А' - вещества-изомеры. Изомеразы - сравнительно немногочисленный класс ферментов, он подразделяется на следующие подклассы в зависимости от типа катализируемой реакции изомеризации: Рацемазы и эпимеразы катализируют взаимопревращение изомеров, содержащих асимметрические атомы углерода. Рацемазами называются ферменты, действующие на субстраты с одним асимметрическим атомом, например, превращающие L-аминокислоты в D-аминокислоты. Одним из таких ферментов является aлaнинрацемаза (аланин-рацемаза. КФ 5.1.1.1), катализирующая реакцию: 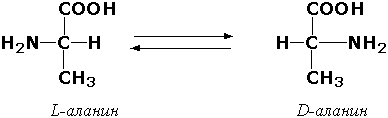 Эпимеразами называются ферменты, действующие на субстраты с несколькими асимметрическими атомами углерода. К таким ферментам относится УДФ-глюкозо-эпимераза (УДФ-глюкоза-4-эпимераза, КФ 5.1.3.2). участвующая в процессах взаимопревращения моносахаридов: 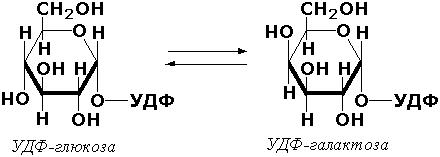 Цис-транс-изомеразы - ферменты, вызывающие изменение геометрической конфигурации относительно двойной связи. Примером такого фермента является малеилацетоацетатизомераза (малеилацетоацетат-цис-транс-изомераза, КФ 5.2.1.2), участвующая в катаболизме фенилаланина и тирозина и переводящая малеилацетоацетат (см. выше реакцию 10) в фумарилацетоацетат: 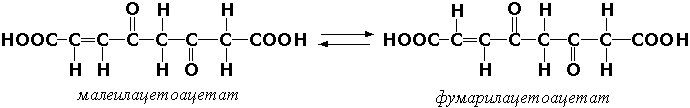 Внутримолекулярные оксидоредуктазы - изомеразы, катализирующие взаимопревращения альдоз и кетоз. При этом происходит окисление СН-ОН-группы с одновременным восстановлением соседней С=О-группы. Так, триозофосфатизомераза (D-глицеральдегид-3-фосфат-кетол-изомераза, КФ 5.3.1.1) катализирует одну из реакций углеводного обмена:  К изомеразам относятся также внутримолекулярные трансферазы, осуществляющие перенос одной группы с одной части молекулы субстрата на другую часть той же молекулы, и внутримолекулярные лиазы, катализирующие реакции дециклизации, а также превращения одного типа кольца в другой. Следует подчеркнуть, что не все биохимические процессы. результатом которых является изомеризация, катализируются изомеразами. Так, изомеризация лимонной кислоты в изопимонную происходит при участии фермента аконитатгидратазы (цитрат (изоцитрат)-гидро-лиаза, КФ 4.2.1.3), катализирующей реакции дегидратации-гидратации с промежуточным образованием цис-аконитовой кислоты: 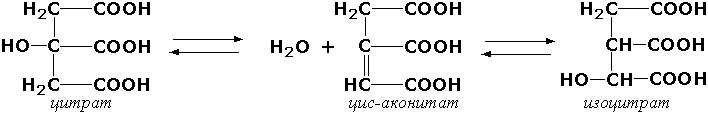 VI. ЛИГАЗЫ Лигазы - класс ферментов, катализирующих синтез органических соединений из активированных за счет распада АТФ (или ГТФ, УТФ, ЦТФ) исходных веществ. Для ферментов этого класса сохраняется также тривиальное название синтетазы. В связи с этим, согласно рекомендациям IUBMB, термин "синтетазы" не должен применяться для ферментов, в действии которых не участвуют нуклеозидтрифосфаты. Реакции, катализируемые лигазами (синтетазами), протекают по схеме: , где А и В - взаимодействующие вещества; А—В - вещество, образующееся в результате взаимодействия. Поскольку в результате действия этих ферментов образуются новые химические связи, подклассы VI класса формируются в зависимости от характера вновь образованных связей. Лигазы, образующие связи углерод-кислород. К ним относится группа ферментов, известных как аминокислота-тРНК-лигазы (аминоацил-тРНК-синтетазы). которые катализируют реакции взаимодействия аминокислот и соответствующих транспортных РНК. В этих реакциях образуются активные формы аминокислот, способные участвовать в процессе синтеза белка на рибосомах. Примером фермента может служить тирозил-тРНК-синтетаза (L-тирозин:тРНК-лигаза (АМФ), КФ 6.1.1.1), участвующая в реакции: 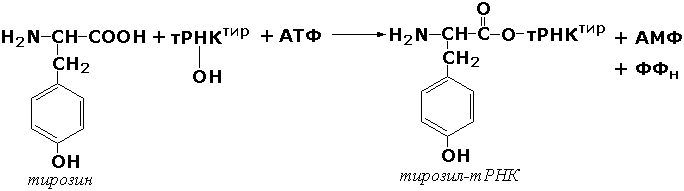 Лигазы, образующие связи углерод-сера. Этот подкласс представлен в первую очередь ферментами, катализирующими образование тиоэфиров жирных кислот с коэнзимом А. При участии этих ферментов синтезируются ацил-КоА - активные формы жирных кислот, способные вступать в различные реакции биосинтеза и распада. Рассмотрим одну из реакций активации жирных кислот, протекающую в присутствии фермента ацил-КоА-синтетазы (карбоновая кислота:коэнзим А-лигаза (АМФ). КФ 6.2.1.2): 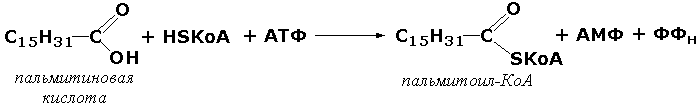 Лигазы, образующие связи углерод-азот, катализируют многочисленные реакции введения азотсодержащих групп в органические соединения. Примером может служить глутаминсинтетаза (L-глутамин:аммиак-γ-лигаза (АДФ), КФ 6.3.1.2). участвующая в обезвреживании токсичного продукта обмена - аммиака - в реакции с глутаминовой кислотой: 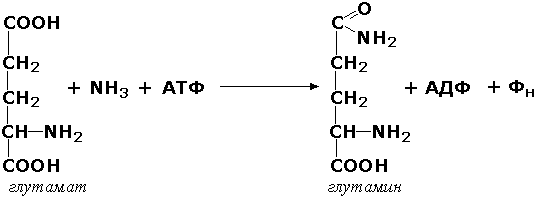 Лигазы, образующие связи углерод-углерод. Из этих ферментов наиболее изучены карбоксилазы, обеспечивающие карбоксилирование ряда соединений, в результате чего происходит удлиннение углеродных цепей. Важнейшим представителем данного класса является пируваткарбоксилаза (пируват:СО2-лигаза (АДФ), КФ 6.4.1.1), ускоряющая реакцию образования оксалоацетата - ключевого соединения цикла трикарбоновых кислот и биосинтеза углеводов: 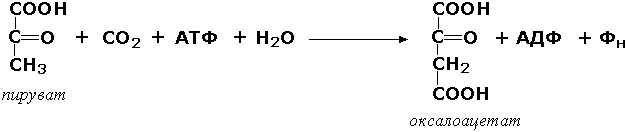 Напомним, что реакции с участием АТФ катализируются не только ферментами VI класса, но и некоторыми ферментами II класса (фосфотрансферазами или киназами). Важно уметь отличать эти типы реакций. Их различие заключается в том, что в трансферазных реакциях АТФ является донором фосфатных групп, поэтому в результате этих реакцию не происходит выделения Н3РО4 (примеры см. выше). Наоборот, в синтетазных реакциях АТФ служит источником энергии, выделяемой при её гидролизе, поэтому одним из продуктов такой реакции будет являться неорганический орто- или пирофосфат. Свойства ферментов. Ферменты сочетают в себе свойства катализаторов и свойства, присущие всем остальным белкам. Как и все белки, ферменты: состоят из многих сотен аминокислотных остатков, соединённых в определённой последовательности при помощи пептидных связей; они имеют глобулярную пространственную структуру, определяющую их функциональные возможности; различаются по молекулярной массе, электрофоретической подвижности, образуют коллоидные растворы. Подобно неорганическим катализаторам, ферменты: не расходуются в процессе реакции, увеличивают скорость как прямой, так и обратной реакции, не изменяют положения равновесия. Белковая природа ферментов обусловливает появление у них ряда свойств, в целом нехарактерных для неорганических катализаторов: олигодинамичность, специфичность, зависимость скорости реакции от температуры, рН среды, концентрации фермента и субстрата, присутствия активаторов и ингибиторов. Олигодинамичность Ферменты обладают высокой эффективностью даже в очень небольших количествах. Такая высокая эффективность объясняется тем, что молекулы ферментов в процессе своей каталитической деятельности непрерывно регенерируют. Типичная молекула фермента может регенерировать миллионы раз в минуту. Надо сказать, что и неорганические катализаторы также способны ускорять превращение такого количества веществ, которое во много раз превышает их собственную массу. Но ни один неорганический катализатор не может сравниться с ферментами по эффективности действия. Примером может служить фермент реннин, вырабатываемый слизистой оболочкой желудка жвачных животных. Одна молекула его за 10 минут при 37°С способна вызывать коагуляцию (створаживание) порядка миллиона молекул казеиногена молока. Другой пример высокой эффективности ферментов даёт каталаза. Одна молекула этого фермента при 0°С расщепляет за секунду около 50 000 молекул пероксида водорода: 2 Н2О2 2 Н2О + О2 Действие каталазы на пероксид водорода заключается в изменении величины энергии активации этой реакции приблизительно от 75 кДж/моль без катализатора до 21 кДж/моль в присутствии фермента. Если же в качестве катализатора этой реакции используется коллоидная платина, то энергия активации составляет всего 50 кДж/моль. Специфичность действия ферментов Важное свойство, отличающее ферменты от неорганических катализаторов – специфичность. Структура активного центра фермента комплементарна структуре его субстрата. Поэтому фермент из всех имеющихся в клетке веществ выбирает и присоединяет только свой субстрат. Для ферментов характерна специфичность не только по отношению к субстрату, но и в отношении пути превращения субстрата. У ферментов различают абсолютную, относительную и стереохимическую специфичность. Абсолютная специфичность – избирательная способность фермента катализировать только единственное из возможных превращений одного субстрата. Это можно объяснить конформационной и электростатической комплементарностью молекул субстрата и фермента. Например, фермент аргиназа катализирует только гидролиз аминокислоты аргинина, фермент уреаза – только расщепление мочевины и не действуют на другие субстраты. Относительная специфичность – избирательная способность фермента катализировать однотипные превращения сходных по строению субстратов. Такие ферменты оказывают воздействие на одинаковые функциональные группы или на один и тот же тип связей в молекулах субстратов. Так, например, разные гидролитические ферменты действуют на определённый тип связей: амилаза – на гликозидные связи; пепсин и трипсин – на пептидные связи; липаза и фосфолипаза – на сложноэфирные связи. Действие этих ферментов распространяется на большое число субстратов, что позволяет организму обойтись малым количеством пищеварительных ферментов - иначе их потребовалось бы намного больше. Стереохимическая (оптическая) специфичность - избирательная способность фермента катализировать превращение только одного из возможных пространственных изомеров субстрата. Так, большинство ферментов млекопитающих катализирует превращение толькл L-изомеров аминокислот, но не D-изомеров. ферменты, участвующие в обмене моносахаридов, наоборот, катализируют превращение только D-, но не L-фосфосахаров. Гликозидазы специфичны не только к моносахаридному фрагменту, но и характеру гликозидной связи. Например, α-амилаза расщепляет α–1,4-гликозидные связи в молекуле крахмала, но не действует на α–1,2-гликозидные связи в молекуле сахарозы. |