биохимия лекции. Энзимология как учение о ферментах. Простые и сложные ферменты
 Скачать 3.9 Mb. Скачать 3.9 Mb.
|

|
Раздел 2.5 Зависимость скорости ферментативной реакции от концентрации субстрата, температуры и рН среды. При изучении влияния какого-либо фактора на скорость ферментативной реакции все прочие факторы должны оставаться неизменными и по возможности иметь оптимальное значение. Мерой скорости ферментативных реакций служит количество субстрата, подвергшегося превращению в единицу времени, или количество образовавшегося продукта. Изменение скорости проводят на начальной стадии реакции, когда продукт ещё практически отсутствует, и обратная реакция не идёт. Кроме того, на начальной стадии реакции концентрация субстрата соответствует его исходному количеству. Зависимость скорости ферментативной реакции (V) от концентрации фермента [Е] (рисунок 2.3). При высокой концентрации субстрата (многократно превышающей концентрацию фермента) и при постоянстве других факторовскорость ферментативной реакции пропорциональна концентрации фермента. Поэтому зная скорость реакции, катализируемой ферментом, можно сделать вывод о его количестве в исследуемом материале.  Рисунок 2.3. Зависимость скорости ферментативной реакции от концентрации фермента Зависимость скорости реакции от концентрации субстрата [S]. График зависимости имеет вид гиперболы (рисунок 2.4). При постоянной концентрации фермента скорость катализируемой реакции возрастает с увеличением концентрации субстрата до максимальной величины Vmax, после чего остаётся постоянной. Это следует объяснить тем, что при высоких концентрациях субстрата все активные центры молекул фермента оказываются связанными с молекулами субстрата. Любое избыточное количество субстрата может соединиться с ферментом лишь после того, как образуется продукт реакции и освободится активный центр. 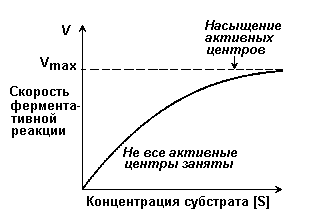 Рисунок 2.4. Зависимость скорости ферментативной реакции от концентрации субстрата. Зависимость скорости реакции от концентрации субстрата может быть выражена уравнением Михаэлиса — Ментен: ,  где V — скорость реакции при концентрации субстрата [S] , Vmax —максимальная скорость и KM —константа Михаэлиса. Константа Михаэлиса равна концентрации субстрата, при которой скорость реакции составляет половину максимальной. Определение KM и Vmax имеет важное практическое значение, так как позволяет количественно описать большинство ферментативных реакций, включая реакции с участием двух и более субстратов. Различные химические вещества, изменяющие активность ферментов, по-разному воздействуют на величины Vmax и KM. Зависимость скорости реакции от t – температуры, при которой протекает реакция (рисунок 2.5), имеет сложный характер. Значение температуры, при котором скорость реакции максимальна, представляет собой температурный оптимум фермента. Температурный оптимум большинства ферментов организма человека приблизительно равен 40°С. Для большинства ферментов оптимальная температура равна или выше тойц температуры, при которой находятся клетки.  Рисунок 2.5. Зависимость скорости ферментативной реакции от температуры. При более низких температурах (0° — 40°С) скорость реакции увеличивается с ростом температуры. При повышении температуры на 10°С скорость ферментативной реакции удваивается (температурный коэффициент Q10 равен 2). Повышение скорости реакции объясняется увеличением кинетической энергии молекул. При дальнейшем повышении температуры происходит разрыв связей, поддерживающих вторичную и третичную структуру фермента, то есть тепловая денатурация. Это сопровождается постепенной потерей каталитической активности. Зависимость скорости реакции от рН среды (рисунок 2.6). При постоянной температуре фермент работает наиболее эффективно в узком интервале рН. Значение рН, при котором скорость реакции максимальна, представляет собой оптимум рН фермента. У большинства ферментов организма человека оптимум рН находится в пределах рН 6 – 8, но есть ферменты, которые активны при значениях рН, лежащих за пределами этого интервала. Изменение рН как в кислую, так и в щелочную сторону от оптимума приводит к изменению степени ионизации кислых и основных групп аминокислот, входящих в состав фермента (например, СООН-группы аспартата и глутамата, NН2-группы лизина и т.д.). Это вызывает изменение конформации фермента, в результате чего изменяется пространственная структура активного центра и снижение его сродства к субстрату. Кроме того, при экстремальных значениях рН происходит денатурация фермента и его инактивация.  Рисунок 2.6. Зависимость скорости ферментативной реакции от рН среды. Следует отметить, что свойственный ферменту оптимум рН не всегда совпадает с рН его непосредственного внутриклеточного окружения. Это позволяет предположить, что среда, в которой находится фермент, в какой-то мере регулирует его активность. Зависимость скорости реакции от присутствия активаторов и ингибиторов. Активаторы повышают скорость ферментативной реакции. Ингибиторы понижают скорость ферментативной реакции. В качестве активаторов ферментов могут выступать неорганические ионы. Предполагают, что эти ионы заставляют молекулы фермента или субстрата принять конформацию, способствующую образованию фермент-субстратного комплекса. Тем самым увеличивается вероятность взаимодействия фермента и субстрата, а следовательно и скорость реакции, катализируемой ферментом. Так, например, активность амилазы слюны повышается в присутствии хлорид-ионов. Раздел 3.2 Особенности структурной организации активного центра фермента. Каким же образом ферменты повышают скорость химических реакций? Представим себе смесь двух веществ. Реакция между этими веществами возможна, но она не пойдёт без затраты некоторого количества энергии. Энергия, необходимая для того, чтобы заставить вещества вступить в реакцию, называется энергией активации [Ea] (рисунок 3.1). Чем больше необходимая энергия активации, тем ниже скорость реакции при данной температуре.  Рисунок 3.1. Энергетические барьеры катализируемой и некатализируемой реакций. В любой совокупности молекул того или иного вещества индивидуальные молекулы при постоянной температуре сильно различаются по количеству содержащейся в них энергии. Лишь небольшая часть их может преодолеть активационный барьер и вступить в реакцию в отсутствие катализатора. Поэтому скорость реакции в таких условиях будет очень низкой. Фермент, соединяясь с субстратом, образует короткоживущий фермент-субстратный комплекс (рисунок 3.2), которому соответствует более низкая энергия активация по сравнению с субстратом в некатализируемой реакции; такой энергией обладает уже значительно больше молекул субстрата. По завершении реакции фермент-субстратный комплекс распадается на продукт (или продукты) и фермент. Фермент по окончании реакции остаётся таким же, как был до неё, и может взаимодействовать с новой молекулой субстрата.  Рисунок 3.2. Образование фермент-субстратного комплекса в ходе катализируемой реакции. Именно таким образом ферменты снижают энергетический барьер реакции: в их присутствии гораздо большее число молекул вступает в реакцию за единицу времени. Активный центр фермента В процессе формирования фермент-субстратного комплекса субстрат присоединяется к специфическому участку на молекуле фермента, который называется активным центром. Активный центр – участок молекулы фермента, который связывает субстраты и от которого зависит специфичность каталитического действия ферментов; активный центр содержит функциональные группы остатков аминокислот и коферментов, пространственно сближенных и определённым образом ориентированных. Несмотря на огромное разнообразие структуры ферментов, их специфичности и механизма действия, существует ряд общих закономерностей формирования активных центров. Во-первых, на активный центр приходится относительно малая часть объёма фермента. Роль остальных аминокислотных остатков, составляющих основную массу фермента, состоит в том, чтобы обеспечить молекуле фермента правильную глобулярную форму. Во-вторых, активный центр – это сложная трёхмерная структура, и в её образовании принимают участие группы, принадлежащие разным частям линейной последовательности аминокислот. Радикалы аминокислот, образующих активный центр, оказываются вблизи друг от друга в результате формирования третичной структуры белка (рисунок 3.3). Поэтому при воздействии факторов, вызывающих денатурацию (нагревание, концентрированные кислоты и щёлочи) утрачивается конформация активного центра и фермент теряет свою активность.  Рисунок 3.3. А. Участие аминокислотных остатков, образующих активный центр фермента, во взаимодействии с субстратом. Б. Положение этих аминокислотных остатков в первичной структуре фермента. В-третьих, активный центр имеет форму узкого углубления или щели, в которую ограничен доступ воде, за исключением тех случаев, когда вода является одним из реагирующих веществ. В этом углублении присутствует несколько полярных аминокислотных остатков, необходимых для связывания субстрата и катализа. В-четвёртых, в составе активного центра можно условно выделить две части: а) контактный или якорный участок, где происходит связывание субстрата в нужной ориентации; б) каталитический участок, обеспечивающий протекание реакции.  Рисунок 3.4. Состав активного центра фермента (на примере химотрипсина). В-пятых, субстраты относительно слабо связываются с ферментами. В связывании и превращении субстрата принимают участие следующие группировки аминокислотных радикалов: полярные заряженные: карбоксильные группы глутамата и аспартата, аминогруппы лизина; гуанидиновые группы аргинина; имидазольные группы гистидина; полярные незаряженные: гидроксильные группы серина и треонина; сульфгидрильные группы цистеина; фенольные группы тирозина; неполярные группы: углеводородные цепи алифатических аминокислот; ароматические кольца фенилаланина и триптофана. У сложных ферментов в формировании активных центров принимают участие также функциональные группы коферментов. В образовании фермент-субстратных комплексов принимают участие те же молекулярные взаимодействия, что и обеспечивают формирование пространственной структуры макромолекул, межклеточные контакты и другие процессы в биологических системах: водородные связи между полярными незаряженными группировками субстрата и фермента; ионные связи между противоположно заряженными группировками субстрата и фермента; гидрофобные взаимодействия между неполярными группировками субстрата и фермента. Эти три основных типа нековалентных связей различаются по своей геометрии, энергии, специфичности. Раздел 3.3 Взаимодействие активного центра с субстратом: модели жёсткого и индуцированного соответствия. Cпецифичность связывания субстрата с ферментом зависит от строго определённого расположения атомов в активном центре. Субстрат входит в активный центр, если он соответствует ему по форме. Существует две модели, описывающие взаимодействие субстрата с активным центром: 1. Модель жёсткого соответствия («ключ – замок»), предложена Э. Фишером в 1890 году. Активный центр считается заранее подогнанным под форму молекулы субстрата (рисунок 3.5). Эта модель не утратила своего значения для понимания некоторых свойств ферментов, например, их способности к строго определённому связыванию двух или большего числа субстратов или для объяснения кинетики насыщения субстратом.  Рисунок 3.5. Взаимодействие субстрата с ферментом согласно модели жёсткого соответствия (Л.Страйер, 1984). 2. Модель индуцированнного соответствия («рука – перчатка»), предложена Кошлендом в 1950-е годы. Согласно этой модели, субстрат вызывает (индуцирует) конформационные изменения фермента, и лишь в результате этих изменений аминокислотные остатки фермента принимают пространственную ориентацию, необходимую для связывания субстрата и катализа (рисунок 3.6). При этом другие аминокислотные остатки могут погрузиться вглубь молекулы фермента.  Рисунок 3.6. Взаимодействие субстрата с ферментом согласно модели индуцированного соответствия (Л.Страйер, 1984). Значение конформационных изменений, возникающих в молекуле фермента в процессе присоединения к ней субстрата можно рассмотреть на примере гексокиназы. Этот фермент катализирует фосфорилирование глюкозы в реакции с АТФ. Присоединение относительно небольшой молекулы глюкозы к активному центру гексокиназы приводит к сближению полипептидных цепей двух субъединиц, которые, как клещи, захватывают молекулу глюкозы (рисунок 3.7). По-видимому, при такой индуцированной подгонке конформации фермента к структуре субстрата молекула глюкозы также деформируется и облегчается её взаимодействие с молекулой АТФ. 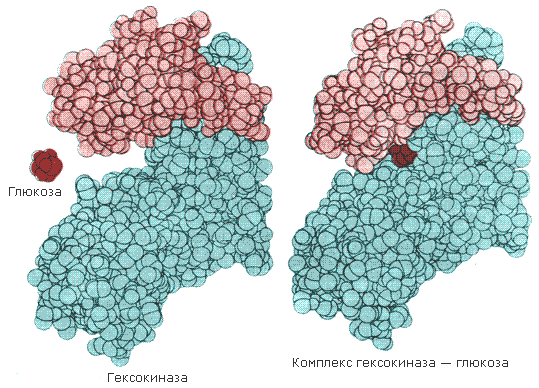 Рисунок 3.7. Индуцированное соответствие при взаимодействии гекокиназы с глюкозой по данным рентгеноструктурного исследования (А.Ленинджер, 1985) Раздел 3.4 Аллостерические ферменты: изменение конформации под действием эффекторов. Виды аллостерической регуляции. Ферменты, осуществляющие в клетке различные метаболические процессы, организованы в виде последовательных цепей или систем, в которых они действуют согласованно. В каждой такой ферментной системе есть хотя бы один фермент, выполняющий роль «дирижера», который задает скорость всей последовательности реакций, так как он катализирует лимитирующую стадию, т. е. самую медленную реакцию, определяющую скорость всего процесса в целом. Такие ферменты-«дирижеры» обладают способностью повышать или понижать свою каталитическую активность в ответ на определенные сигналы. Благодаря действию подобных ферментов скорость каждой последовательности метаболических реакций постоянно изменяется, почти мгновенно приспосабливаясь к изменяющимся потребностям клетки в энергии и играющих роль строительных блоков молекулах, необходимых для роста и обновления клеток. В большинстве случаев фермент-«дирижер» катализирует первую реакцию такой последовательности. Остальные же ферменты просто подчиняются указаниям «дирижера»; катализируемые ими реакции ускоряются лишь при поступлении достаточного количества субстратов, образующихся в качестве продуктов предшествующих реакций. Такие ферменты-«дирижёры», активность которых изменяется под действием молекулярных сигналов различных типов, называются регуляторными. Существуют два основных класса регуляторных ферментов: аллостерические, т.е. ферменты, регулируемые нековалентно связанными с ними модуляторами, и ферменты, регулируемые путём их ковалентной модификации. Многие ферменты, обладающие олигомерной структурой (хотя и не все), являются аллостерическими белками, способными изменять своё сродство к субстрату. Аллостерические ферменты, как правило, катализируют начальные реакции в многостадийных путях химических превращений в клетке. Аллостерические ферменты отличаются от остальных ферментов тем, что не подчиняются классической кинетике Михаэлиса – Ментен. Зависимость скорости реакции от концентрации субстрата у таких ферментов имеет вид S-образной кривой. Наряду с активным центром такие ферменты содержат по меньшей мере один аллостерический центр (регуляторный центр). Аллостерический центр - участок молекулы фермента, способный присоединять определённые молекулы (эффекторы или модуляторы). Аллостерический центр специфичен по отношению к своему эффектору подобно тому, как активный центр специфичен по отношению к своему субстрату. Между аллостерическим центром одной из субъединиц фермента и аллостерическим эффектором могут возникать нековалентные взаимодействия (водородные, ионные и гидрофобные). Это приводит к обратимому изменению конформации остальных субъединиц молекулы фермента, в том числе изменению конформации активного центра. В результате активность фермента снижается или повышается. 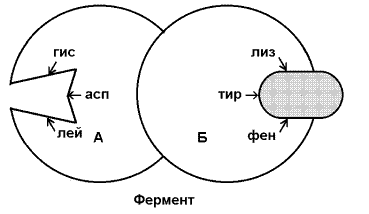 Рисунок 3.8. Схема взаимодействия аллостерического фермента и его эффектора. Аллостерический фермент состоит из двух субъединиц: А - каталитической, включающей активный центр, и Б - регуляторной, в состав которой входит аллостерический центр. Присоединение эффектора к аллостерическому центру приводит к изменению конформации активного центра. Аллостерические эффекторы бывают двух типов – активаторы и ингибиторы. Аллостерические активаторы способствуют переходу фермента из Т-конформации с низким сродством к субстрату в R-конформацию с высоким сродством к субстрату, аллостерические ингибиторы – наоборот. Если после присоединения эффектора сродство активного центра фермента к субстрату повышается, то эффектор называется аллостерическим активатором, если сродство понижается, то эффектор называется аллостерическим ингибитором. Различают гомотропную и гетеротропную аллостерическую регуляцию. В случае гомотропной регуляции эффектором является субстрат. У таких ферментов аллостерический центр по своей конформации совпадает с активным, а роль аллостерического эффектора фермента выполняет молекула субстрата. Взаимодействие субстрата с активным центром одной из субъединиц аллостерического фермента повышает сродство остальных субъединиц к субстрату. Это напоминает связывание молекулы гемоглобина с кислородом. В случае гетеротропной регуляции эффектор отличается от субстрата и аллостерический центр не совпадает с активным центром. Примером может служить регуляция биосинтеза пиримидиновых нуклеотидов. 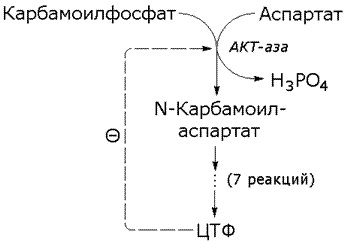 Рисунок 3.9. Схема биосинтеза пиримидиновых нуклеотидов. Начальную реакцию этого метаболического пути катализирует фермент аспартат-карбамоилтрансфераза (АКТ-аза). Конечный продукт цепи реакций – цитидинтрифосфат (ЦТФ) является аллостерическим ингибитором АКТ-азы. При увеличении концентрации ЦТФ сродство фермента к субстратам снижается, хотя максимальная скорость реакции (Vmax) остаётся неизменной. График зависимости активности от концентрации при этом смещается вправо. В этом случае Vmax может быть достигнута при более высокой концентрации субстрата (аспартата). Ингибирующее действие ЦТФ может быть снято добавлением АТФ (субстратом промежуточных реакций биосинтеза). 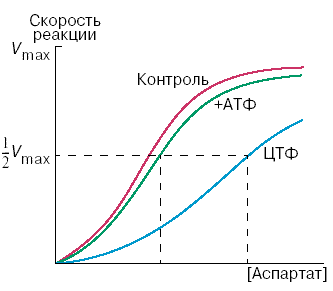 Рисунок 3.10. Регуляция аспартат-карбамоилтрансферазы ЦТФ и АТФ. Таким образом, при накоплении ЦТФ в клетке скорость синтеза пиримидиновых нуклеотидов снижается и повышается при снижении концентрации ЦТФ. Так фермент обеспечивает постоянное присутствие в клетке нужных количеств цитидинтрифосфата.. |