биохимия лекции. Энзимология как учение о ферментах. Простые и сложные ферменты
 Скачать 3.9 Mb. Скачать 3.9 Mb.
|

|
Раздел 8.3 Разобщение дыхания и фосфорилирования. 8.3.1. Перенос электронов в дыхательной цепи не во всех случаях протекает сопряжённо с фосфорилированием АДФ. Состояние, при котором окисление субстратов в дыхательной цепи происходит, но АТФ при этом не образуется, называется свободным (нефосфорилирующим) окислением. Энергия, выделяемая при окислении, рассеивается в виде теплоты. В физиологических условиях свободное окисление может служить одним из механизмов терморегуляции. В организме человека и некоторых животных имеется особая ткань – бурый жир, содержащий митохондрии, приспособленные для выработки теплоты. Много бурого жира у новорождённых, в последующие периоды жизни его количество уменьшается. В митохондриях бурого жира содержание дыхательных ферментов значительно выше, чем ферментов, осуществляющих фосфорилирование АДФ, поэтому в них преобладают процессы свободного окисления. Разобщение процессов окисления и фосфорилирования в митохондриях может иметь место при некоторых патологических состояниях. Основными симптомами таких состояний могут быть быстрая утомляемость, повышенная температура тела, снижение массы тела, несмотря на повышенный аппетит, учащение дыхания и сердцебиения. 8.3.2. Разобщение процессов окисления и фосфорилирования может быть вызвано действием ряда веществ, как природных, так и синтетических. Механизм действия этих веществ заключается в том, что они являются переносчиками протонов через мембрану. Вещества, разобщающие окисление и фосфорилирование, можно разделить на протонофоры и ионофоры. Протонофоры представляют собой слабые гидрофобные органические кислоты, которые в форме аниона (R-COO-) связывают протоны в межмембранном пространстве, диффундируют через мембрану и диссоциируют в матриксе с образованием протонов. К этой группе относятся, например, свободные жирные кислоты, гормоны щитовидной железы, салицилаты, дикумарол, 2,4-динитрофенол (см. рисунок 8.6). 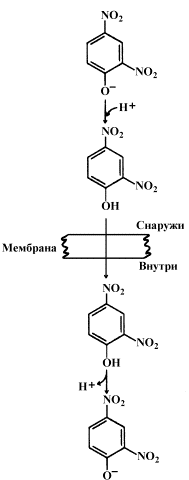 Рисунок 8.6. Механизм действия 2,4-динитрофенола. Ионофоры (валиномицин, нигерицин, грамицидин) способны встраиваться в мембрану, образуя канал, по которому могут перемещаться протоны и другие одновалентные катионы - Na+ или K+ (рисунок 8.7). В результате снимается протонный потенциал и нарушается синтез АТФ.  Рисунок 8.7. Валиномицин облегчает проникновение в клетку ионов Н+. 8.4.1. Микросомальное окисление является одним из этапов биотрансформации – обезвреживания неполярных (нерастворимых в воде) соединений как эндогенного происхождения, так и чужеродных для организма (ксенобиотиков). Эндогенные субстраты - холестерол, стероидные гормоны, ненасыщенные жирные кислоты, витамин D3. Экзогенные субстраты - лекарственные вещества. В результате окисления субстратов повышается их растворимость в воде, скорость выведения из организма. Биотрансформация лекарственных веществ, как правило, снижает их токсичность. 8.4.2. Ферментная система микросомального окисления встроена в мембраны эндоплазматического ретикулума клетки (ЭПР, рисунок 8.8). 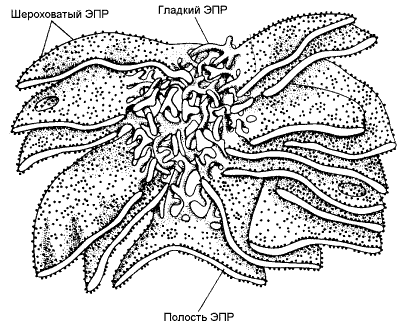 Рисунок 8.8. Структура эндоплазматического ретикулума (источник: Альбертс Б. и соавт., Молекулярная биология клетки, 1994). Она представляет собой короткую цепь переноса водорода и включает несколько последовательно расположенных в мембране белков-ферментов (рисунок 8.9).  Рисунок 8.9. Схема монооксигеназной цепи окисления ЭПР. Источником электронов и протонов в этой цепи является восстановленный кофермент НАДФН, который образуется в реакциях пентозофосфатного пути окисления глюкозы. Промежуточным акцептором Н+ и е— служит флавопротеин (ФлПр), содержащий кофермент ФАД. Конечное звено в цепи микросомального окисления - цитохром Р-450. Это - сложный белок, хромопротеин, в качестве простетической группы содержит гем. Цитохром Р-450 является монооксигеназой, то есть ферментом, включающим один из атомов молекулярного кислорода в окисляемое вещество. Поэтому цепь реакций микросомального окисления называют также монооксигеназной цепью. Цитохром Р-450 выполняет две функции. Он связывает окисляемый субстрат и активирует молекулярный кислород, облегчая их взаимодействие друг с другом. Реакция, катализируемая цитохромом Р-450, называется реакцией гидроксилирования, так как образующийся продукт содержит ОН-группу (рисунок 8.10). 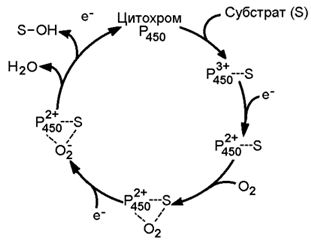 Рисунок 8.10. Механизм реакции гидроксилирования субстрата при участии цитохрома Р450. В отличие от митохондриальной дыхательной цепи, при переносе электронов в монооксигеназной цепи не происходит аккумулирования энергии в виде АТФ. Поэтому микросомальное окисление является свободным окислением. 8.4.3. Основные факторы, влияющие на активность монооксигеназной системы печени, которые следует учитывать при выборе дозировки лекарственных веществ: а) возраст – в детском возрасте и у пожилых людей активность ферментов микросомального окисления ниже, чем у людей среднего возраста; б) пол – мужские половые гормоны повышают скорость реакций гидроксилирования субстратов в печени, женские половые гормоны, наоборот, понижают; в) характер питания – при недостаточном поступлении белков с пищей в печени снижается синтез ферментов, участвующих в микросомальном окислении; г) влияние лекарственных веществ и ядов – некоторые лекарственные препараты (например, фенобарбитал и другие барбитураты, некоторые антибиотики) являются индукторами микросомального окисления и ускоряют процессы гидроксилирования субстратов; другие вещества (угарный газ СО, амизил, дезипрамил) снижают скорость микросомального окисления. Раздел 9.1 Классификация и функции углеводов. 9.1.1. Углеводы - полигидроксикарбонильные соединения и их производные, Их характерным признаком является наличие альдегидной (-СН=О) или кетонной (>C=O) групп и не менее 2 гидроксильных (-ОН) групп. 9.1.2. По структуре углеводы разделяют на моносахариды, олигосахариды и полисахариды. Моносахариды – наиболее простые углеводы, не подвергающиеся гидролизу. В зависимости от наличия альдегидной или кетонной группы различают альдозы (например, глюкоза, галактоза, рибоза, глицеральдегид) и кетозы (например, фруктоза, рибулоза, диоксиацетон). Олигосахариды - углеводы, содержащие от 2 до 10 моносахаридных остатков, соединенных, при помощи гликозидных связей. В зависимости от количества моносахаридных остатков различают дисахариды (содержат 2 остатка, например, лактоза, сахароза, мальтоза), трисахариды (содержат 3 остатка) и.т.д. Полисахариды - углеводы, содержащие более 10 моносахаридных остатков, соединенных при помощи гликозидных связей. Если полисахарид состоит из одинаковых моносахаридных остатков, то это гомополисахарид (крахмал, гликоген, целлюлоза). Если полисахарид состоит из разных моносахаридных остатков, то это гетерополисахарид (гиалуроновая кислота, хондроитинсерная кислота, гепарин). Формулы важнейших углеводов представлены на рисунке 9.1.       Рисунок 9.1. Формулы важнейших углеводов. 9.1.3. Функции углеводов. Углеводы выполняют в организме следующие функции: 1. Энергетическая. Углеводы служат источником энергии. За счет их окисления удовлетворяется примерно половина всей потребности человека в энергии. При окислении 1 г углеводов выделяется около 16,9 кДж энергии. 2. Резервная. Крахмал и гликоген представляют собой форму хранения питательных веществ, выполняя функцию временного депо глюкозы. 3. Структурная. Целлюлоза и другие полисахариды растений образуют прочный остов; в комплексе с белками и липидами они входят в состав биомембран всех клеток. 4. Защитная. Кислые гетерополисахариды выполняют роль биологического смазочного материала, выстилая трущиеся поверхности суставов, слизистой пищеварительных путей, носа, бронхов, трахеи и др. 5. Антикоагулянтная. Гепарин обладает важными биологическими свойствами, в частности препятствует свёртыванию крови. 6. Углеводы являются источником углерода, который необходим для синтеза белков, нуклеиновых кислот, липидов и других соединений. 9.1.4. Источником углеводов для организма служат углеводы пищи (крахмал, сахароза, лактоза, глюкоза). Глюкоза может синтезироваться в организме из аминокислот, глицерина, пирувата и лактата (глюконеогенез). аздел 9.2 Переваривание углеводов в желудочно-кишечном тракте. 9.2.1. Процесс переваривания углеводов представлен на схеме (рисунок 9.2).  Рисунок 9.2. Переваривание углеводов в желудочно-кишечном тракте. Гидролиз крахмала начинается в ротовой полости. В слюне содержится фермент амилаза, частично расщепляющая крахмал. Основное место переваривания крахмала - тонкий кишечник. Туда поступает амилаза сока поджелудочной железы. Продуктом действия амилазы является мальтоза. Мальтоза далее расщепляется с помощью мальтазы до глюкозы, дисахарид лактоза (содержащаяся в молоке) расщепляется с помощью лактазы до глюкозы и галактозы. Дисахарид сахароза (содержащаяся в пищевом сахаре) расщепляется с помощью сахаразы до глюкозы и фруктозы. 9.2.2. Продукты полного переваривания углеводов - глюкоза, галактоза и фруктоза - через клетки кишечника поступают в кровь путем облегченной диффузии и активного транспорта. 9.2.3. Основные пути метаболизма углеводов в тканях. После всасывания из желудочно-кишечного тракта моносахариды поступают через систему портальной вены в печень. В гепатоцитах происходит превращение галактозы и фруктозы в глюкозу. Таким образом, глюкоза является основным моносахаридом, который поступает в общий кровоток после прохождения углеводов через печень. Содержание глюкозы в крови здорового человека составляет 3,33 – 5,55 ммоль/л. Глюкоза поглощается из крови клетками всех тканей и органов. 9.2.4. Избыток глюкозы может откладываться в клетках в виде гликогена – резервного полисахарида с разветвлённой структурой. Окисление глюкозы может происходить дихотомическим и апотомическим путём. Дихотиомическое окисление может происходить без участия кислорода (до молочной кислоты) и при участии кислорода (до СО2 и Н2О). Промежуточные продукты окисления глюкозы, кроме того, могут быть использованы для синтеза глицерола, жирных кислот, аминокислот и нуклеотидов. В клетках различных тканей интенсивность протекания перечисленных путей обмена глюкозы может быть различной. Эти различия определяются прежде всего функциональными особенностями этих тканей. Раздел 9.3 Анаэробное окисление глюкозы. 9.3.1. Гликолиз – это ферментативный распад глюкозы до молочной кислоты (лактата). Гликолиз протекает в тканях без потребления кислорода. В анаэробных условиях гликолиз – единственный процесс, поставляющий АТФ, так как окислительное фосфорилирование в этих условиях не функционирует. Гликолиз протекает в цитоплазме клеток организма. Этот процесс катализируется одиннадцатью ферментами. Условно можно разделить гликолиз на две стадии.  Рисунок 9.3. Реакции первой стадии гликолиза. 9.3.2. Первая стадия гликолиза включает реакции превращения молекулы глюкозы в две молекулы фосфотриоз. Эта стадия сопровождается затратой молекул АТФ. Начальной реакцией глюкозы в клетке является её фосфорилирование в результате взаимодействия с АТФ (рисунок 9.3, реакция 1). Эта реакция в условиях клетки протекает только в одном направлении. Биологическая роль реакции фосфорилирования глюкозы заключается в том, что глюкозо-6-фосфат, в отличие от свободной глюкозы, не может проникать через клеточную мембрану обратно в кровь. В большинстве тканей реакцию фосфорилирования глюкозы катализирует фермент гексокиназа, которая обладает высоким сродством к глюкозе, способна также фосфорилировать фруктозу и маннозу и ингибируется избытком глюкозо-6-фосфата. В клетках печени, кроме того, есть фермент глюкокиназа, которая имеет низкое сродство к глюкозе, не ингибируется глюкозо-6-фосфатом и не участвует в фосфорилировании других моносахаридов. В следующей реакции глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат (рисунок 9.3, реакция 2). Продукт реакции изомеризации подвергается повторному фосфорилированию за счёт АТФ (рисунок 9.3, реакция 3). Эта реакция – наиболее медленно протекающая реакция гликолиза и, подобно фосфорилированию глюкозы, необратима. Фермент – фосфофруктокиназа – является аллостерическим, активируется АДФ и АМФ, ингибируется цитратом и высокой концентрацией АТФ. На следующем этапе фруктозо-1,6-дифосфат подвергается расщеплению на две фосфотриозы (рисунок 9.3, реакция 4). Таким образом, химическое соединение, содержащее 6 углеродных атомов, превращается в два, содержащих по 3 атома углерода. Поэтому гликолиз называют дихотомическим путём превращения глюкозы (от слова дихотомия – рассечение на две части). Далее происходит изомеризация триозофосфатов (рисунок 9.3, реакция 5). В этой реакции диоксиацетонфосфат переходит в глицеральдегид-3-фосфат. Таким образом, в первой стадии гликолиза молекула глюкозы превращается в две молекулы глицеральдегид-3-фосфата.  Рисунок 9.4. Реакции второй стадии гликолиза. 9.9.3. Вторая стадия гликолиза включает реакции превращения двух молекул глицеральдегид-3-фосфата в две молекулы лактата. На этой стадии гликолиза происходит синтез молекул АТФ. Глицеральдегид-3-фосфат подвергается дегидрированию при участии НАД-зависимой дегидрогеназы. В этой реакции происходит потребление неорганического фосфата, который включается в состав продукта реакции, содержащего макроэргическую фосфатную связь (рисунок 9.4, реакция 6). 1,3-Дифосфоглицерат вступает в реакцию первого субстратного фосфорилирования, т.е. не сопряжённого с переносом электронов в дыхательной цепи. В этой реакции осуществляется синтез молекулы АТФ в результате переноса фосфатной группы вместе с макроэргической связью на молекулу АДФ (рисунок 9.4, реакция 7). Раздел 9.4 Особенности обмена сахарозы и лактозы. 9.4.1. Сахароза (её формула приводится на рисунке 9.1) в организм человека поступает с пищевым сахаром. Первая стадия катаболизма сахарозы происходит в желудочно-кишечном тракте под действием фермента сахаразы. Вторая стадия катаболизма (образование пирувата) идёт в цитоплазме клеток печени при участии внутриклеточных ферментов (рисунок 9.5). 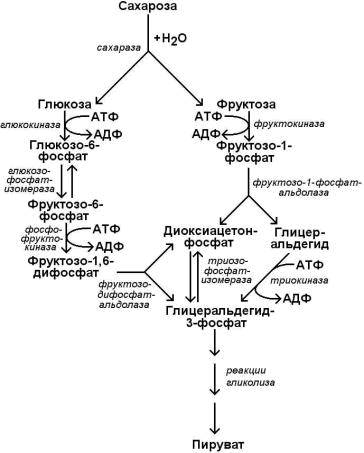 Рисунок 9.5. Схема катаболизма сахарозы. В печени имеется фермент фруктокиназа, который катализирует фосфорилирование фруктозы по 1-му атому углерода, реакция происходит с затратой молекулы АТФ. Образующийся фруктозо-1-фосфат расщепляется далее на диоксиацетонфосфат и глицеральдегид. Оба эти продукта переходят в глицеральдегид-3-фосфат – промежуточный продукт, дальнейшие превращения которого в процессе гликолиза описаны в предыдущем параграфе. При недостатке фруктокиназы нарушается усвоение фруктозы в организме, в крови увеличивается концентрация этого моносахарида и он выводится с мочой. 9.4.2. Лактоза (её формула приводится на рисунке 9.1) поступает в организм с молоком и молочными продуктами. Первая стадия катаболизма лактозы происходит в желудочно-кишечном тракте под действием фермента лактазы. Вторая стадия катаболизма (образование пирувата) идёт в цитоплазме клеток печени при участии внутриклеточных ферментов (рисунок 9.6).  Рисунок 9.6. Схема катаболизма лактозы. Обмен галактозы начинается с превращения её в галактозо-1-фосфат. Эта реакция катализируется галактокиназой и идёт с использованием энергии АТФ. В следующей реакции при участии гексозофосфат-уридилтрансферазы галактозо-1-фосфат превращается в глюкозо-6-фосфат, одновременно образуется УДФ-галактоза. Образовавшийся глюкозо-1-фосфат переходит в глюкозо-6-фосфат и подвергается превращениям, описанным в предыдущем параграфе. УДФ-галактоза подвергается эпимеризации с образованием УДФ-глюкозы. Известны врождённые дефекты ферментов обмена лактозы. Врождённый дефект фермента лактазы приводит к развитию непереносимости лактозы. Употребление в пищу молока сопровождается диспептическими явлениями (рвота, диаррея, метеоризм). Другое наследственное заболевание – галактоземия – возникает при недостатке гексозофосфат-уридил-трансферазы. Для этого заболевания характерно повышение уровня галактозы в крови, катаракта, умственная отсталость. В следующей реакции происходит внутримолекулярное перемещение фосфатной группы 3-фосфоглицерата ко 2-му углеродному атому (рисунок 4, реакция 8). Тем самым облегчается последующее отщепление молекулы воды, которое приводит к появлению в продукте реакции макроэргической фосфатной связи (рисунок 9.4, реакция 9). Фосфоенолпируват (ФЕП) вступает в реакцию второго субстратного фосфорилирования, в ходе которого образуется молекула АТФ. В отличие от первого субстратного фосфорилирования, данная реакция является необратимой в условиях клетки. Фермент пируваткиназа аллостерически ингибируется АТФ и фруктозо-1,6-дифосфатом (рисунок 9.4, реакция 10). В заключительной реакции гликолиза происходит использование НАДН, образовавшегося при дегидрировании глицеральдегид-3-фосфата (см. реакцию 6). При участии НАД-зависимой лактатдегидрогеназы пируват восстанавливается в молочную кислоту (рисунок 9.4, реакция 11). Фермент существует в пяти изоферментных формах, отличающихся сродством к субстрату и распределением в тканях. Таким образом, в процессе гликолиза в клетке не накапливается НАДН. Это значит, что гликолиз может протекать без участия кислорода (который является конечным акцептором электронов, передаваемых НАДН в дыхательную цепь). При подсчёте энергетического баланса гликолиза следует учитывать, что каждая из реакций второй стадии этого метаболического пути повторяется дважды. Таким образом, в первой стадии было затрачено 2 молекулы АТФ, а во второй стадии путём субстратного фосфорилирования образовалось 2х2 = 4 молекулы АТФ; следовательно при окислении одной молекулы глюкозы в клетке накапливается 2 молекулы АТФ. Раздел 9.5 Аэробный дихотомический путь окисления глюкозы. 9.5.1. Аэробным называется окисление биологических субстратов с выделением энергии, протекающее при использовании кислорода в качестве конечного акцептора водорода в дыхательной цепи. В качестве доноров водорода выступают восстановленные формы коферментов (НАДН, ФАДН2 и НАДФН), образующиеся в промежуточных реакциях окисления субстратов. Аэробное дихотомическое окисление глюкозы является основным путём катаболизма глюкозы в организме человека и может происходить во всех органах и тканях. В результате этих реакций глюкоза расщепляется до СО2 и Н2О, а выделяющаяся энергия аккумулируется в АТФ. В этом процессе можно условно выделить три стадии: превращение глюкозы в 2 молекулы пирувата в цитоплазме клеток (специфический путь распада глюкозы); окислительное декарбоксилирование пирувата с образованием ацетил-КоА в митохондриях; окисление ацетил-КоА в цикле Кребса в митохондриях. 9.5.2. На каждом этапе процесса происходит образование восстановленных форм коферментов, которые окисляются ферментными комплексами дыхательной цепи с образованием АТФ путём окислительного фосфорилирования. Коферменты, образующиеся на второй и третьей стадиях аэробного окисления глюкозы, подвергаются непосредственному окислению в митохондриях. В то же время НАДН, образующийся в цитоплазме в реакциях первой стадии аэробного окисления, не способен проникать через митохондриальную мембрану. Перенос водорода с цитоплазматического НАДН в митохондрии происходит при помощи специальных челночных циклов, основным из которых является малат-аспартатный челночный механизм. Цитоплазматический НАДН восстанавливает оксалоацетат в малат, который проникает в митохондрию, где окисляется, восстанавливая митохондриальный НАД; в цитоплазму оксалоацетат возвращается в виде аспартата (рисунок 9.7). 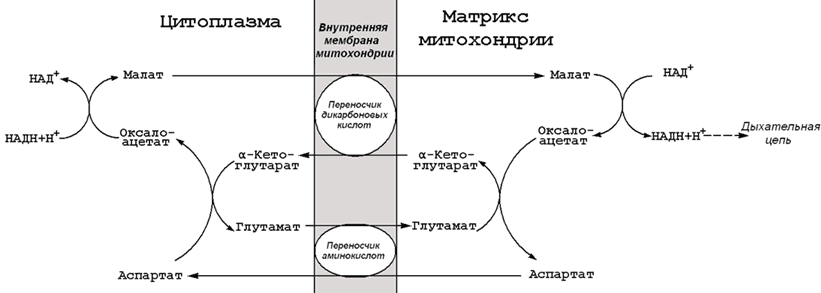 Рисунок 9.7. Малат-аспартатный челночный механизм. Продукция АТФ в реакциях аэробного дихотомического окисления происходит также в трёх реакциях субстратного фосфорилирования – две из них в гликолизе, третья в цикле Кребса на уровне сукцинил-КоА. Полный энергетический баланс аэробного окисления глюкозы представлен на рисунке 9.8.  Рисунок 9.8. Энергетический баланс аэробного окисления глюкозы. |