Общая. Эпителиальные ткани
 Скачать 4.65 Mb. Скачать 4.65 Mb.
|

|
Фиброкласты – клетки дифферона фиброцитов, специализированные на функции разрушения межклеточного вещества соединительной ткани, которая резко преобладает над их синтетической и секреторной активностью. В их цитоплазме выявляются многочисленные вакуоли, содержащие литические ферменты и коллагеновые фибриллы на различных стадиях лизиса. Эти клетки обеспечивают перестройку и инволюцию соединительной ткани Функции фибробластов: Образования межклеточного матрикса (в основном этим занимаются фибробласты) коллаген и эластин, образующие волокна гликозаминогликаны и протеогликаны, формирующие основное вещество Поддержания нормального гомеостаза межклеточного вещества (в основном этим занимаются фиброциты) Секретируют цитокины, регулирующие кроветворение: колониестимулирующие факторы макрофагов и гранулоцитов Участвуют в заживлении ран Вопрос 26. Макрофаги (гистиоциты) и плазматические клетки (СМ и ЭМ), участие в иммунных реакциях. Макрофаги (гистиоциты) - вторые по численности (после фибробластов) клетки рыхлой волокнистой соединительной ткани. Они принадлежат к линии потомков стволовой клетки крови и непосредственно образуются из моноцитов после их миграции в соединительную ткань из просвета кровеносных сосудов. В соединительной ткани макрофаги располагаются поодиночке или группами. Эти клетки очень многочисленны в собственной пластинке слизистых оболочек, а также в серозных оболочках. Они могут пребывать в одном из двух взаимообратимых состояний: Покоящихся клеток, обладающих низкой функциональной активностью Блуждающих клеток с высокой функциональной активностью Покоящиеся гистиоциты трудно идентифицировать на светооптическом уровне. Они имеют вид мелких уплощенных клеток удлиненной или отростчатой формы с четкими контурами, прикрепленных к коллагеновым волокнам. Эти клетки характеризуются небольшим земным ядром и плотной цитоплазмой со слабо развитыми органеллами. При активации, происходящей под действием микроорганизмов или их продуктов, а также ряда цитокинов, клетки в покое могут превращаться в блуждающие. Блуждающие (активные) гистиоциты обладают высокой подвижностью, изменчивой (отростчатой, реже округлой) формой с неровными, но обычно четко выявляемыми краями. Их ядро светлее, чем в покоящихся клетках, но темнее, чем в фибробластах; в нем может выявляться ядрышко. Цитоплазма содержит многочисленные лизосомы и развитые элементы цитосклета, которые концентрируются в области псевдоподий; другие органеллы развиты умеренно. В иммунных реакциях макрофаги выполняют роль АПК, т. е. участвуют в иммунных реакциях посредством захвата, процессинга антигена, предоставления комплекса [эпитоп + МНС класса 2] Т-хелперам и их активация. Также они синтезируют цитокины, стимулирующие пролиферацию лимфоцитов и вовлечение их в иммунную реакцию. Плазматические клетки (плазмоциты) и их предшественники Влимфоциты, находящиеся на различных этапах преобразования в плазмоциты - в небольших количествах постоянно содержатся в различных участках рыхлой волокнистой соединительной ткани. Это неподвижные или очень слабо подвижные, короткоживущие. Плазматические клетки имеют сравнительно мелкие размеры (диаметр 9-20 мкм, в среднем - 14 мкм), круглую или овальную форму. Ядро - округлое, расположено эксцентрично, содержит крупные глыбки гетерохроматина, которые располагаются в виде радиальных тяжей ("спиц колеса"). Ядрышко крупное, лежит в центре ядра или эксцентрично. В отличие от других клеток, интенсивно вырабатывающих белок, продукты синтетической деятельности плазматических клеток (иммуноглобулины) в норме не накапливаются в цитоплазме в секреторных гранулах, а по мере образования транспортируется мелкими пузырьками к плазмолемме, где непрерывно выделяется механизмом экзоцитоза. Функция плазматических клеток заключается в обеспечении гуморального иммунитета путем выработки антител. Вопрос 27. Биосинтез коллагена. Уровни структурной организации коллагеновых и эластических волокон. Типы коллагена, зоны локализации в организме. Молекулы коллагена построены из триплетов – трех пептидных цепочек (а(альфа)-цепочки) – проколлагена, свивающихся ещё в клетке в единую спираль. Этот процесс проходит в клетке в несколько этапов: Синтез а-цепи проколлагена Гидроксилирование (присоединение –ОН групп) части остатков пролина и лизина Гликолизирование (присоединение остатков сахаров) определенных остатков гидроксилизина Объединение а-цепей проколлагена и образование молекулы коллагена. Последующий процессинг происходит вне клетки. Выделяют 4 уровня структурной организации коллагеновых волокон: Молекулярный – триплет проколлагена. Надмолекулярный – соединённые в длину и в ширину с помощью водородных связей молекулы тропоколлагена (триплеты без концевых пептидов). Сначала образуются протофибриллы, а 5-6 протофибрилл, скрепленных между собой связями, составляют микрофибриллы. Фибриллярный уровень – объединённые в фибриллы гликозаминогликанами (также синтезированные фибробластами) микрофибриллы. Волоконный уровень – объединенные в волокно протеогликанами фибриллы. 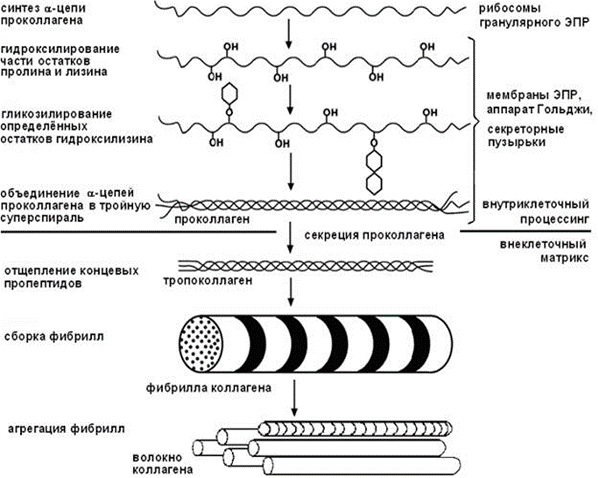 Уровни организации эластического волокна: Молекулярный уровень – глобула (не фибрилла) диаметром 2,8нм Эластиновые протофибриллы толщиной 3—3,5 нм – соединённые в цепочки глобулы Микрофибриллы – Эластиновые протофибриллы объединяются друг с другом в сочетании с гликопротеином (фибриллином) Волокно – Объединённые в длину и ширину микрофибриллы. Содержат около 90 % аморфного компонента эластических белков (эластина) в центре, а по периферии — микрофибриллы.  Вопрос 28. Плотная волокнистая соединительная ткань, ее разновидности и функции. Строение сухожилия. Плотная волокнистая соединительная ткань (ПВСТ) характеризуются относительно большим количеством плотно расположенных волокон и незначительным количеством клеточных элементов и основного аморфного вещества между ними. Основная функция ПВСТ – обеспечение механической прочности. В зависимости от характера расположения волокнистых структур эта ткань подразделяется на плотную неоформленную и плотную оформленную соединительную ткань.
Сухожилие (tendo). Оно состоит из толстых, плотно лежащих параллельных пучков коллагеновых волокон. Между этими пучками располагаются фиброциты и небольшое количество фибробластов и основного аморфного вещества. Каждый пучок коллагеновых волокон, отделенный от соседнего слоем фиброцитов, называется пучком первого порядка. Несколько пучков первого порядка, окруженных тонкими прослойками рыхлой волокнистой соединительной ткани, составляют пучки второго порядка. Прослойки рыхлой волокнистой соединительной ткани, разделяющие пучки второго порядка, называются эндотенонием. Из пучков второго порядка слагаются пучки третьего порядка, разделенные более толстыми прослойками рыхлой соединительной ткани — перитенонием (сухожилие – tendo). Иногда пучком третьего порядка является само сухожилие. В крупных сухожилиях могут быть и пучки четвертого порядка. В перитенонии и эндотенонии проходят кровеносные сосуды, питающие сухожилие, нервы и проприоцептивные нервные окончания, посылающие в центральную нервную систему сигналы о состояции натяжения ткани сухожилий. Некоторые сухожилия (самые крупные и подвижные) в местах прикрепления к костям заключены во влагалища, построенные из двух волокнистых соединительнотканных оболочек, между которыми находится жидкость (смазка), богатая гиалуроновой кислотой. Вопрос 29. Хрящевая ткань. Классификация хрящевых тканей. Дифферон хрящевой ткани. Строение и функции надхрящницы. Зональность строения хряща (на примере гиалиновой хрящевой ткани). Хрящевые ткани входят в состав органов дыхательной системы, суставов, межпозвоночных дисков итд, состоят из клеток — хондроцитов и хондробластов и большого количества межклеточного гидрофильного вещества, отличающегося упругостью. От 50 до 70 % сухого вещества хрящевой ткани составляет коллаген, волокна которого преимущественно направлены против действия механических сил (то есть если на хрящ нагрузка идёт сверху, то и волокна в этом хряще изначально занимают вертикальное положение), что помогает аммортизации. Различают три вида хрящевой ткани: гиалиновую, эластическую, волокнистую. Такое подразделение хрящевых тканей основано на структурно-функциональных особенностях строения их межклеточного вещества, степени содержания и соотношения коллагеновых и эластических волокон. 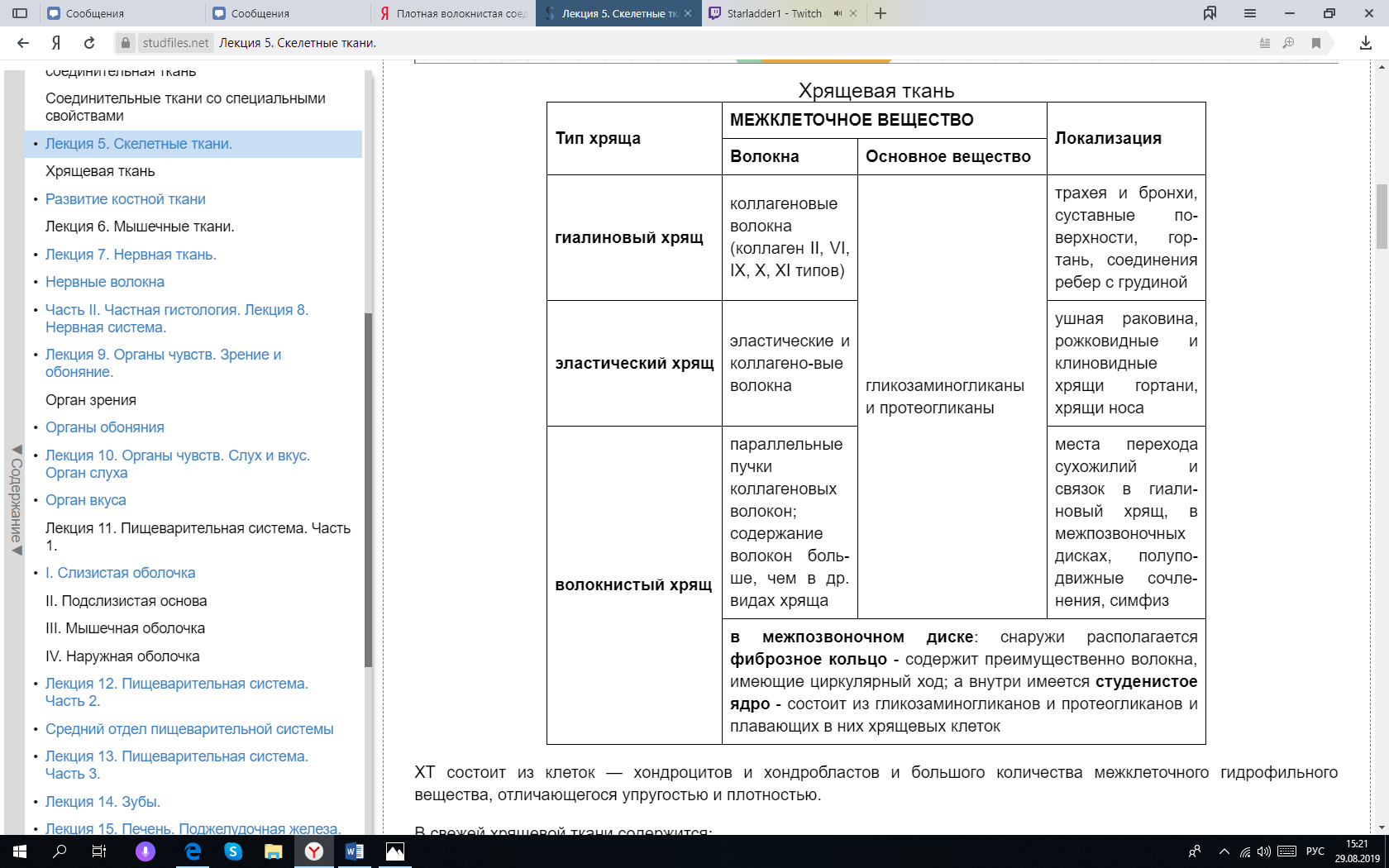 Клетки хрящевых тканей представлены хондробластическим дифференом: Стволовая клетка. Полустволовая клетка. Хондробласты. Молодые клетки, располагаются в глубоких слоях надхрящницы по одиночке, не образуя изогенные группы. Основная функция хондробластов - выработка органической части межклеточного вещества. Хондробласты способны к размножению и в последующем превращаются в хондроциты. В целом, хондробласты обеспечивают поверхностный рост хряща со стороны надхрящницы. Хондроциты. Основные клетки хрящевой ткани, располагаются в более глубоких слоях хряща в полостях - лакунах. Хондроциты могут делиться митозом, при этом дочерние клетки не расходятся, образуют изогенные группы. Основная функция хондроцитов - выработка органической части межклеточного вещества хрящевой ткани. Рост хряща за счет деления хондроцитов и выработки ими межклеточного вещества обеспечивает внутренний рост хряща. Хондрокласты. Они являются разрушителями поврежденной или изношенной части хряща. В норме в хряще их нет или представлены в очень маленьком количестве. Надхрящница - это слой соединительной ткани, покрывающий поверхность хряща. В надхрящнице выделяют наружный фиброзный (из плотной неоформленной СТ с большим количеством кровеносных сосудов) слой и внутренний клеточный слой, содержащее большое количество стволовых, полустволовых клеток и хондробластов. Так как сам хрящ не имеет сосудов внутри, но его межклеточное вещество обладает высокой гидрофильностью, питание происходит диффузно за счёт сосудов надхрящницы. В хряще (как органе) выявляются 2 нерезко разграниченных слоя (зоны), в пределах каждого из которых хрящевая ткань характеризуется рядом морфологических, биохимических и функциональных особенностей. Зона молодого хряща располагается в виде сравнительно тонкого слоя непосредственно над надхрящницей. Она состоит из уплощённых молодых хондробластов, лежащих поодиночке параллельно поверхности хряща, которые окружены гомогенным оксифильным матриксом. Зона зрелого хряща образует его основную массу и располагается глубже предыдущей. В области плавного перехода из зоны молодого хряща хондроциты становятся более округлыми, а ещё глубже они располагаются в виде изогенных групп. Вопрос 30. Гистогенез хрящевых тканей. Механизм интерстициального и аппозиционного роста. Источником развития хрящевых тканей является мезенхима. В первой стадии в некоторых участках тела зародыша, где образуется хрящ, клетки мезенхимы теряют свои отростки, усиленно размножаются и, плотно прилегая друг к другу, создают определенное напряжение – тургор. Такие участки называют хондрогенными зачатками, или хондрогенными островками. Находящиеся в их составе стволовые клетки дифференцируются в хондробласты. В их цитоплазме сначала увеличивается количество свободных рибосом, затем появляются участки гранулярной эндоплазматической сети. В следующей стадии – образования первичной хрящевой ткани, клетки центрального участка (первичные хондробласты) округляются, увеличиваются в размере, в их цитоплазме развивается гранулярная эндоплазматическая сеть, с участием которой происходят синтез и секреция фибриллярных белков (коллагена). В дальнейшем – в стадии дифференцировки хрящевой ткани – хондроциты приобретают способность синтезировать гликозаминогликаны, кроме упомянутых ранее фибриллярных белков, главным образом сульфатированные (хондроитинсульфаты), связанные с неколлагеновыми белками (протеогликаны). По периферии хрящевой закладки, на границе с мезенхимой формируется надхрящница – оболочка, покрывающая развивающийся хрящ снаружи и состоящая из наружного волокнистого и внутреннего хондрогенного (камбиального) слоев. В хондрогенной зоне клетки интенсивно делятся, дифференцируются в хондробласты, которые сохраняют способность к синтезу ДНК, размножению, а также к синтезу компонентов межклеточного вещества. В процессе секреции продуктов синтеза хондрогенного слоя надхрящницы и наслаивания на уже имеющийся хрящ по его периферии сами клетки «замуровываются» в продукты своей деятельности. Так происходит рост хряща способом наложения, или аппозиционный рост. Хрящевые клетки, лежащие в центре молодого развивающегося хряща, сохраняют способность в течение некоторого времени делиться митотически, оставаясь в одной лакуне (изогенные группы клеток), и вырабатывать межклеточное вещество. За счет увеличения количества этих клеток происходит увеличение массы хряща изнутри, что называется интерстициальным ростом. Интерстициальный рост наблюдается в эмбриогенезе, а также при регенерации хрящевой ткани. Вопрос 31. Костные ткани. Классификация. Диффероны костной ткани. (СМ и ЭМ). Характеристика матрикса. Механизм минерализации и резорбции. Костные ткани — это специализированный тип соединительной ткани с высокой минерализацией межклеточного органического вещества, содержащего около 70 % неорганических соединений, главным образом фосфатов кальция. Органическое вещество — матрикс костной ткани — представлено в основном белками коллагенового типа и липидами. Органические и неорганические компоненты в сочетании друг с другом определяют механические свойства — способность сопротивляться растяжению, сжатию и др. Из всех разновидностей соединительных тканей костная ткань обладает наиболее выраженными опорной, механической, защитной функциями для внутренних органов, а также является депо солей кальция, фосфора и др. Существует два основных типа костной ткани: ретикулофиброзная (грубоволокнистая) и пластинчатая. Они различаются по структурным и физическим свойствам, которые обусловлены главным образом строением межклеточного вещества. К костной ткани относятся также дентин и цемент зуба. Костный дифферон: Стволовые клетки – морфологически не идентифицируются Полустволовые клетки (преостеобласты) – морфологически не идентифицируются Остеобласты (разновидность фибробластов) – это молодые клетки, создающие костную ткань. В сформировавшейся кости они встречаются только в глубоких слоях надкостницы и в местах регенерации костной ткани после ее травмы. Форма остеобластов бывает различной: кубической, пирамидальной или угловатой. Размер их тела около 15—20 мкм. Ядро округлой или овальной формы. В цитоплазме остеобластов хорошо развиты гранулярная эндоплазматическая сеть, митохондрии и аппарат Гольджи. (они создают кость, поэтому создают белки кости, обычная белоксинтезирующая клетка) Остеоциты – это преобладающие по количеству дефинитивные клетки костной ткани, утратившие способность к делению. Они имеют отростчатую форму, компактное, относительно крупное ядро и слабобазофильную цитоплазму. Органеллы развиты слабо. Также в кости присутствуют остеокласты. Они не имеют отношения к костному дифферону, а являются разновидностью макрофагов, то есть развиваются из СКК в ККМ. Один остеокласт может разрушить столько кости, сколько создают 100 остеобластов за это же время. Функции остеобластов и остеокластов взаимосвязаны и коррелируют с участием гормонов, простагландинов, функциональной нагрузкой, витаминами итд. Остеокласты – довольно крупные клетки, они содержат от 3 до нескольких десятков ядер. Цитоплазма слабобазофильна, иногда оксифильна, содержит мало органелл. На одной из сторон клетки имеются выросты цитоплазмы. Характеристика матрикса. Органический компонент матрикса составляет 30 % от массы кости. Из этих 30% 95 % приходится на коллаген I типа, остальные 5 % представлены протеогликанами и белками, специфическими для кости (остеонектин и остеокальцин). Минерализация и резорбция. Несмотря на высокую степень минерализации, в костных тканях происходят постоянное обновление входящих в их состав веществ, постоянное разрушение и созидание, адаптивные перестройки к изменяющимся условиям функционирования. Остеобласты минерализуют кость, а остеокласты участвуют в процессе резорбции. В остеобластах находится щелочная фосфатаза, в остеокластах кислая фосфатаза. Минерализация происходит в 2 этапа: Остеобласты синтезируют коллаген первого типа, снабжены рецепторами паратгормона, их основная функция – формирование органического остеоида и его минерализация. В процессе минерализации также участвуют неколлагеновые кальций-связывающие белки остеобластов В зоне минерализации усиливаются окислительные процессы, распадается гликоген, синтезируется необходимое количество АТФ. Кроме того, в остеобластах увеличивается количество цитрата, необходимого для синтеза аморфного фосфата кальция. А потом цепочка химических реакций (настолько сложных и непонятных, что даже писать нет смысла) и формируются кристаллы гидроксилапатита вдоль коллагеновых волокон. Основную остеолитическую функцию в зонах ремоделирования кости выполняют остеокласты. Остеокласты перемещаются в эти зоны и образуют в участках резорбируемой кости активный слой. Они выделяют коллагеназу и кислую фосфатазу, лизируя минерализованный остеоид и растворяя кристаллы гидроксиапатита. Вопрос 32. Строение трубчатой кости. Надкостница. Собственное вещество кости. Гормональная регуляция. Трубчатая кость как орган в основном построена из пластинчатой костной ткани, кроме бугорков. Строение диафиза. Компактное вещество, образующее диафиз кости, состоит из костных пластинок, толщина которых колеблется от 4 до 12— 15 мкм. Костные пластинки располагаются в определенном порядке, образуя сложные образования (гаверсовы системы). В диафизе различают три слоя: наружный слой общих пластинок, средний, образованный концентрически напластованными вокруг сосудов костными пластинками — остеонами и называемый остеонным слоем, и внутренний слой общих пластинок. Наружные общие (генеральные) пластинки не образуют полных колец вокруг диафиза кости, перекрываются на поверхности следующими слоями пластинок. В наружных общих пластинках залегают прободающие (фолькмановы) каналы, по которым из надкостницы внутрь кости входят сосуды. Внутренние общие пластинки хорошо развиты только там, где компактное вещество кости непосредственно граничит с костномозговой полостью. В тех же местах, где компактное вещество переходит в губчатое, его внутренние общие пластинки продолжаются в пластинки перекладин губчатого вещества. В среднем слое костные пластинки располагаются главным образом в остеонах, формируя остеонные пластинки, а также вставочные пластинки (бывшие остеоны), лежащие между остеонами. Остеоны (гаверсовы системы) являются структурными единицами компактного вещества трубчатой кости. Они представляют собой цилиндрические образования, состоящие из костных пластинок, как бы вставленных друг в друга. В костных пластинках и между ними располагаются тела костных клеток и их отростки, замурованные в костном межклеточном веществе. Каждый остеон отграничен от соседних остеонов так называемой спайной линией, образованной основным веществом, цементирующим их. В центральном канале (гаверсов канал) остеона проходят кровеносные сосуды с сопровождающей их соединительной тканью и остеогенными клетками. Вставочные пластинки заполняют пространства между остеонами и являются остатками ранее существовавших остеонов, разрушенных в процессе перестройки кости Строение эпифиза. Эпифиз представляет собой губчатое вещество. Губчатое вещество – относительно легкое, мягкие ткани составляют в нем 75% объема. Оно состоит из трехмерной сети анастомозирующих трабекул (дуг, арок), разделенных межтрабекулярными пространствами, содержащими костный мозг. Такое строение обеспечивает не только большую площадь поверхности (порядка 10 м2), на которой осуществляются метаболические процессы, происходящие в кости, но и придает высокую механическую прочность при относительно небольшой массе. Наиболее толстые и мощные трабекулы располагаются в направлении действия максимальных механических нагрузок. Надкостница покрывает кость снаружи и прочно прикреплена к ней толстыми пучками коллагеновых прободающих волокон, которые проникают и вплетаются в слой наружных общих пластинок кости. В надкостнице имеются два слоя: Наружный слой надкостницы образован плотной волокнистой неоформленной соединительной тканью, в которой преобладают волокна, идущие параллельно поверхности кости. Надкостница без резких границ переходит в участки прикрепления связок и мышц. Внутренний слой надкостницы (у взрослых различим слабо, потому что нужен в основном для остеогенеза) состоит из рыхлой волокнистой соединительной ткани, в которой располагаются плоские веретеновидные клетки - покоящиеся остеобласты и их предшественники (преостеобласты). Функции надкостницы: Трофическая - надкостница обеспечивает питание кости, поскольку она содержит сосуды, которые (вместе с нервами) проникают из нее в кость через особые питательные отверстия. Регенераторная - обусловлена наличием в ее внутреннем слое камбиальных элементов Механическая, опорная - надхрящница обеспечивает механическую связь кости с другими структурами (сухожилиями, связками, мышцами), прикрепляющимся к ней. Собственное вещество кости – это и есть пластинчатая и губчатая костная ткань. Гормональная регуляция. При избытке гормона околощитовидной железы — паратирина — наблюдаются повышение активности остеокластов и резорбция кости. Тирокальцитонин, вырабатываемый С-клетками щитовидной железы, действует диаметрально противоположно, понижая функцию остеокластов, имеющих к этому гормону рецепторы. При гипофункции щитовидной железы замедляется рост длинных трубчатых костей в результате подавления активности остеобластов и торможения процесса оссификации. Регенерация кости в этом случае происходит слабо и неполноценно. В случае тестикулярной недоразвитости или препубертатной кастрации задерживается окостенение метаэпифизарной пластинки, вследствие чего руки и ноги у такого индивидуума становятся непропорционально длинными. При недостатке эстрогенов после наступления климактерического периода у женщин иногда развивается остеопороз. При раннем половом созревании намечается остановка роста из-за преждевременного диафизо-эпифизарного сращения костей. Определенную позитивную роль в росте костей имеет соматотропный гормон аденогипофиза, который стимулирует пропорциональное развитие скелета в молодом (юношеском) возрасте и непропорциональное (акромегалия) у взрослых. Вопрос 33. Гистогенез костных тканей. Прямой и непрямой остеогенез. Строение эпифизарной хрящевой пластинки роста. Прямой остеогенез. Такой способ остеогенеза характерен для развития грубоволокнистой костной ткани при образовании плоских костей, например, покровных костей черепа. Этот процесс наблюдается в основном в течение первого месяца внутриутробного развития и характеризуется образованием сначала первичной «перепончатой», остеоидной костной ткани с последующей импрегнацией (отложением) солей кальция, фосфора и др. в межклеточном веществе. В первой стадии – образование скелетогенного островка – в местах развития будущей кости происходят очаговое размножение мезенхимных клеток и васкуляризация скелетогенного островка. Во второй стадии, заключающейся в дифференцировке клеток островков, образуется оксифильное межклеточное вещество с коллагеновыми фибриллами — органическая матрица костной ткани. Разрастающиеся волокна раздвигают клетки, которые, не теряя своих отростков, остаются связанными друг с другом. В основном веществе появляются мукопротеиды (оссеомукоид), цементирующие волокна в одну прочную массу. Некоторые клетки, дифференцирующиеся в остеоциты, уже в этой стадии могут оказаться включенными в толщу волокнистой массы. Другие, располагающиеся по поверхности, дифференцируются в остеобласты. В течение некоторого времени остеобласты располагаются по одну сторону волокнистой массы, но вскоре коллагеновые волокна появляются и с других сторон, отделяя остеобласты друг от друга. Постепенно эти клетки оказываются «замурованными» в межклеточном веществе, теряют способность размножаться и превращаются в остеоциты. В то же время из окружающей мезенхимы образуются новые генерации остеобластов, которые наращивают кость снаружи (аппозиционный рост). Третья стадия — кальцификация межклеточного вещества. При этом остеобласты выделяют фермент щелочную фосфатазу, расщепляющую содержащиеся в периферической крови глицерофосфаты на углеводные соединения (сахара) и фосфорную кислоту. Последняя вступает в реакцию с солями кальция. Непрямой остеогистогенез. На 2-м месяце эмбрионального развития в местах будущих трубчатых костей закладывается из мезенхимы хрящевой зачаток, который очень быстро принимает форму будущей кости (хрящевая модель). Зачаток состоит из эмбрионального гиалинового хряща, покрытого надхрящницей. Развитие кости на месте хряща начинается в области диафиза. Образованию перихондральной костной манжетки предшествует разрастание кровеносных сосудов с дифференцировкой в надхрящнице, прилежащей к средней части диафиза, остеобластов, образующих в виде манжетки сначала ретикулофиброзную костную ткань (первичный центр окостенения), затем заменяющуюся на пластинчатую. Образование костной манжетки нарушает питание хряща. Вследствие этого в центре диафизарной части хрящевого зачатка возникают дистрофические изменения. Рост хряща в этом месте прекращается. Удлинение перихондральной костной манжетки сопровождается расширением зоны деструкции хряща и появлением остеокластов, которые очищают пути для врастающих в модель трубчатой кости кровеносных сосудов и остеобластов. Это приводит к появлению очагов эндохондрального окостенения (вторичные центры окостенения). В связи с продолжающимся ростом соседних неизмененных дистальных отделов диафиза хондроциты на границе эпифиза и диафиза собираются в колонки, направление которых совпадает с длинной осью будущей кости. Таким образом, в колонке хондроцитов имеются два противоположно направленных процесса — размножение и рост в дистальных отделах диафиза и дистрофические процессы в его проксимальном отделе (то есть растёт в сторону эпифиза, а окостеневает в сторону противоположную от эпифиза). С момента разрастания сосудистой сети и появления остеобластов надхрящница перестраивается, превращаясь в надкостницу. В дальнейшем кровеносные сосуды с окружающей их мезенхимой, остеогенными клетками и остеокластами врастают через отверстия костной манжетки и входят в соприкосновение с обызвествленным хрящом. Под влиянием ферментов, выделяемых остеокластами, происходит растворение (хондролиз) обызвествленного межклеточного вещества. Диафизарный хрящ разрушается, в нем возникают удлиненные пространства, в которых «поселяются» остеоциты, образующие на поверхности оставшихся участков обызвествленного хряща костную ткань. В теченииие всего периода роста кость увеличивается как в длину, так и в ширину. Рост трубчатой кости в длину обеспечивается наличием метаэпифизарной хрящевой пластинки роста, в которой проявляются два противоположных гистогенетических процесса. Один — это разрушение эпифизарной пластинки, а другой, противоположный ему, — непрестанное пополнение хрящевой ткани путем новообразования клеток. Однако со временем процессы разрушения клеток начинают преобладать над процессами новообразования, вследствие чего хрящевая пластинка истончается и исчезает. В метаэпифизарном хряще различают пограничную зону, зону столбчатых клеток и зону пузырчатых клеток. Пограничная зона, расположенная вблизи эпифиза, состоит из округлых и овальных клеток и единичных изогенных групп, которые обеспечивают связь хрящевой пластинки с костью эпифиза. В полостях между костью и хрящом находятся кровеносные капилляры, обеспечивающие питанием клетки глубжележащих зон хрящевой пластинки. Зона столбчатых клеток содержит активно размножающиеся клетки, которые формируют колонки, расположенные по оси кости, и обеспечивают ее рост и длину. Проксимальные концы колонок состоят из созревающих, дифференцирующихся хрящевых клеток. Зона пузырчатых клеток характеризуется гидратацией и разрушением хондроцитов с последующим эндохондральным окостенением. Дистальный отдел этой зоны граничит с диафизом, откуда в нее проникают остеогенные клетки и кровеносные капилляры. Вопрос 34. Мышечное волокно. Световая, поляризационная и электронная микроскопия. Миофибрилла. Саркомер, строение, формула саркомера. Мышечное волокно – структурная единица скелетной мышечной ткани. Состоит из миосимпласта и миосателитоцитов, которые покрыты общей базальной мембраной. Длина всего волокна может измеряться сантиметрами при толщине 50-100мкм. СМ. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. У полюсов ядер располагаются органеллы общего значения. Миофибриллы заполняют основную часть миосимпласта и расположены продольно. Сильное развитие имеет аЭПС, которая называется саркоплазматической сетью (продольно окружают миофибрилы своими петлями) и является депо Ca2+. Поляризационная микроскопия. Тут видим деление на саркомеры. Саркомер — структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные А-диски и изотропные I-диски). ЭМ. С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, он перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются) Миофибриллы имеют вид нитей диаметром 1-2 мкм и длиной, сопоставимой с протяженностью волокна. Они обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что А- и I- диски одних миофибрилл точно совпадают с аналогичными дисками других, обусловливая поперечную исчерченность всего волокна. Структурно-функциональной единицей миофибриллы является саркомер. Структура саркомера представлена упорядоченной системой толстых и тонких белковых нитей (миофиламентов). Толстые нити – миозиновые. Связаны с М-линией. Протеолитическими ферментами миозин расщепляется на две фракции - легкий меромиозин ("стержень" молекулы миозина) и тяжелый меромиозин (участки головок и шейки, связывающие их со стержневой частью). Миозин головок обладает АТФазной активностью Тонкие нити содержат сократимый белок актин (F-актин, т.е. фибриллярный) и два регуляторных белка - тропонин и тропомиозин. Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуя длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Тропонин состоит из трех субъединиц: ТnС - связывающей кальций, ТnТ - прикрепляющейся к тропомиозину, и TnI - ингибирующей связывание миозина с актином. 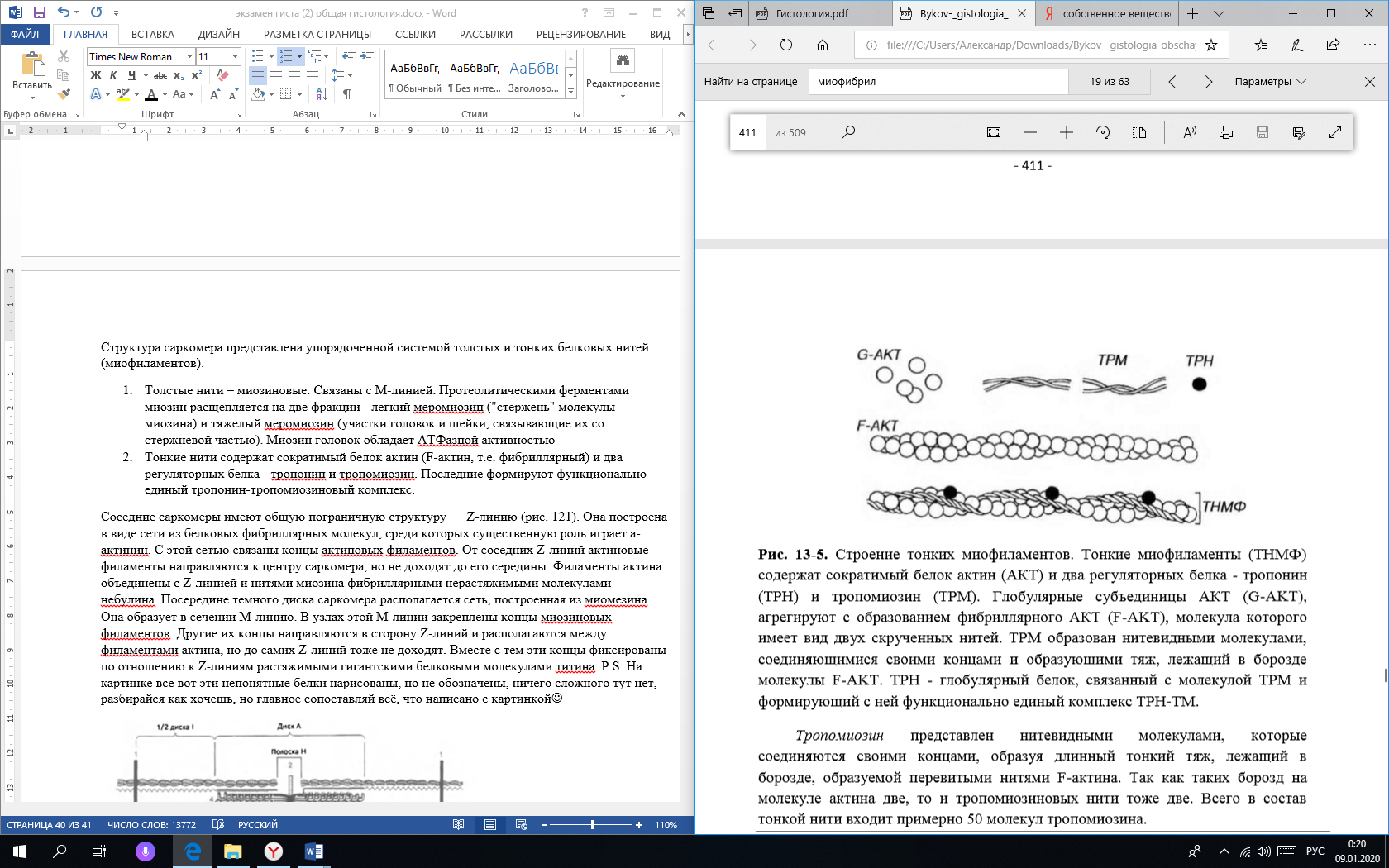 Соседние саркомеры имеют общую пограничную структуру — Z-линию. Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет а-актинин. С этой сетью связаны концы актиновых филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию. В узлах этой М-линии закреплены концы миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.!!!ФОРМУЛА:1/2I+A+1/2I=саркомер!!! P.S. На картинке все вот эти непонятные белки нарисованы, но не обозначены, ничего сложного тут нет, разбирайся как хочешь, но главное сопоставляй всё, что написано с картинкой 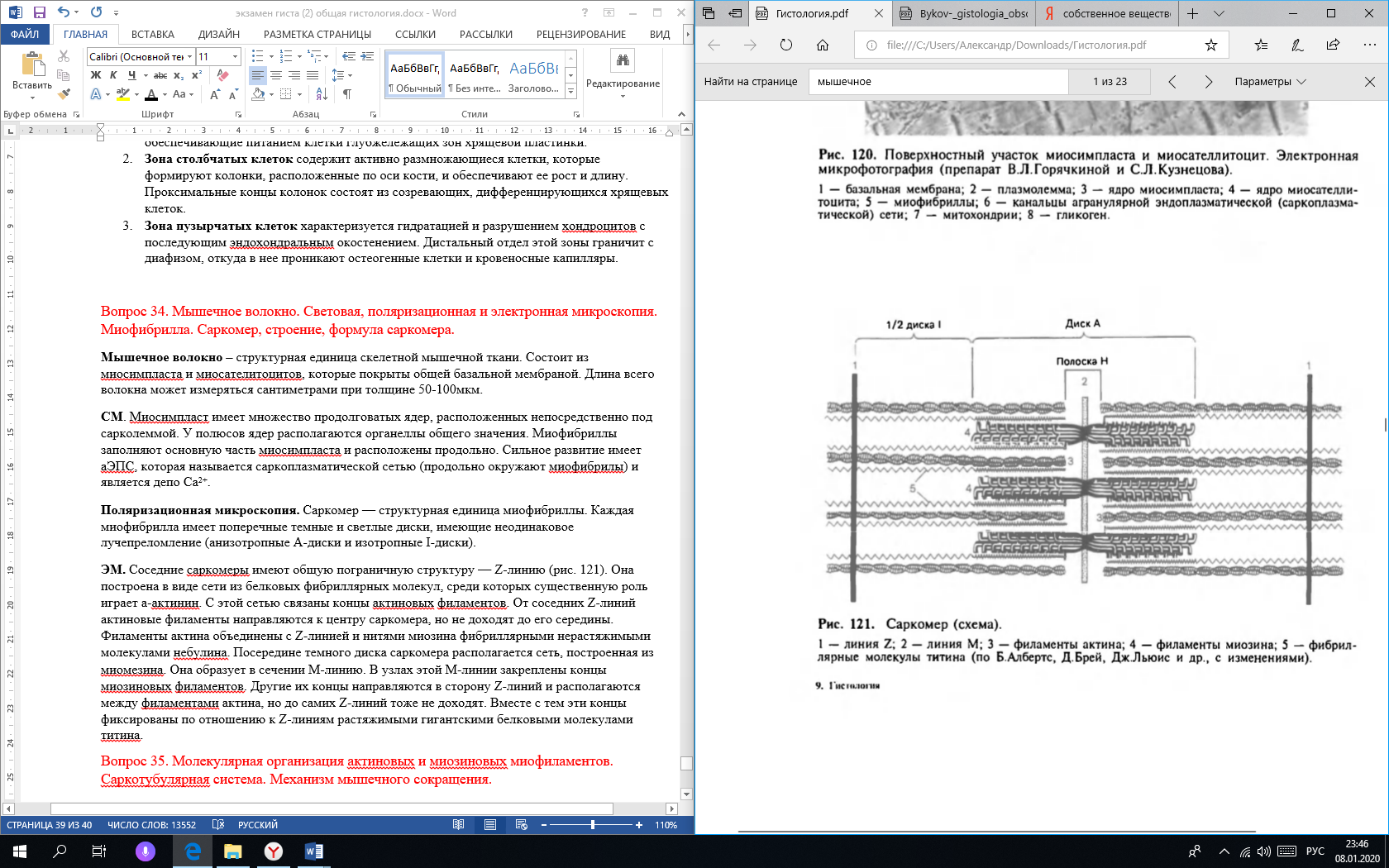 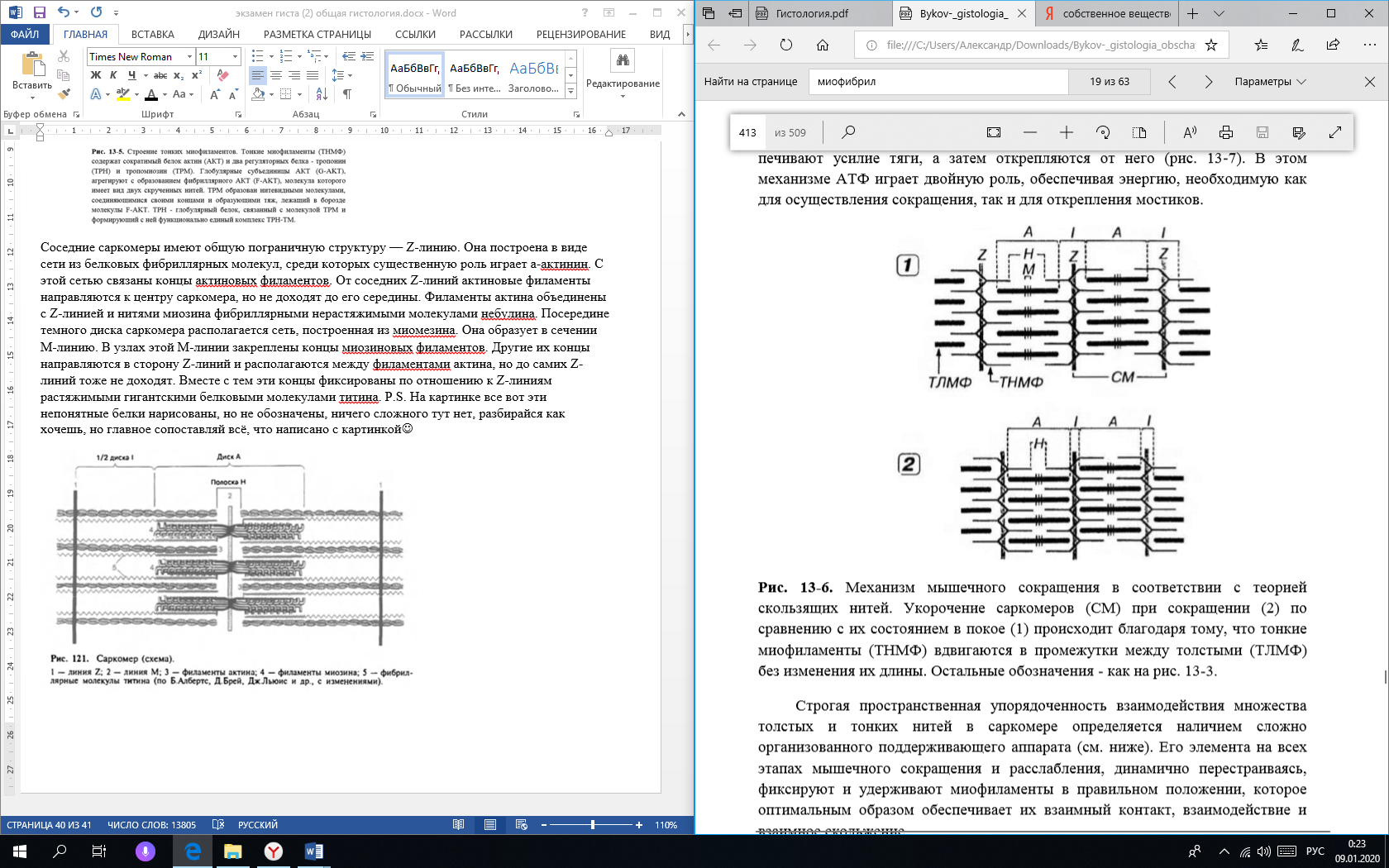 Вопрос 35. Молекулярная организация актиновых и миозиновых миофиламентов. Саркотубулярная система. Механизм мышечного сокращения. Актиновые миофиламенты содержат сократимый белок актин (на него приходится 20% белков миофибриллы) и два регуляторных белка - тропонин (около 2%) и тропомиозин (около 7%). Последние формируют функционально единый тропонин-тропомиозиновый комплекс. Актин в мономерной форме представлен полярными глобулярными субъединицами диаметром 4-5 нм (G-актин). G-актин агрегирует с образованием полимерного фибриллярного актина (F-актина), молекула которого имеет вид двух скрученных нитей толщиной 7 нм и вариабельной длины. Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуя длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Так как таких борозд на молекуле актина две, то и тропомиозиновых нити тоже две. Всего в состав тонкой нити входит примерно 50 молекул тропомиозина. Тропонин представляет собой глобулярный белок, каждая его молекула располагается на тропомиозиновой молекуле вблизи ее конца. Тропонин состоит из трех субъединиц: ТnС - связывающей кальций, ТnТ - прикрепляющейся к тропомиозину, и TnI - ингибирующей связывание миозина с актином. Миозиновые миофиламенты образованы упорядоченно упакованными молекулами фибриллярного белка миозина, на который приходится около 54% всех белков миофибриллы. Молекула миозина имеет вид нити длиной 150 нм и толщиной 2 нм. На одном из концов эта молекула содержит две округлые головки длиной около 20 нм и шириной около 4 нм. Миозин головок обладает АТФазной активностью. Протеолитическими ферментами миозин расщепляется на две фракции - легкий меромиозин ("стержень" молекулы миозина) и тяжелый меромиозин (участки головок и шейки, связывающие их со стержневой частью). Аппарат передачи возбуждения (саркотубулярная система) необходим для того, чтобы распространяющаяся по сарколемме волна деполяризации могла вызвать срабатывание сократительного аппарата миофибрилл. В мышечном волокне связь между возбуждением и сокращением выполняют две специализированные мембранные системы - саркоплазматическая сеть и поперечные (Т-) трубочки, образующие функционально единую саркоту-булярную систему. Саркоплазматическая сеть - система уплощенных, вытянутых и анастомозирующих мембранных трубочек и мешочков, которая окружает каждый саркомер миофибриллы наподобие муфты. Саркоплазматическая сеть обладает выраженной способностью депонировать и выделять ионы кальция. Ее мембрана содержит высокие концентрации интегральных белков, являющихся кальциевыми насосами, а на внутренней поверхности находится белок кальсеквестрин, связывающий ионы Са2+. Поперечные (Т-) трубочки представляют собой впячивания сарколеммы, отходящие от нее под прямым углом к оси волокна и расположенные у млекопитающих вблизи границы I- и А- дисков. Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что тонкие нити вдвигаются в промежутки между толстыми без изменения их длины. Мышечное сокращение вызывается резким повышением концентрации ионов Са2+ в области миофиламентов и включает несколько этапов: Связывание ионов Са2+ с тропонином и освобождение активных центров на молекуле актина. Ионы Са2+ связываются с ТnС-субъединицами тропонина на тонких филаментах. При этом тропонин изменяет свою конформацию, смещает молекулы тропомиозина и открывает активные центры. Связывание миозина и актина (формирование поперечных мостиков). Миозиновые головки связываются с активными центрами на молекуле актина, формируя мостики, расположенные перпендикулярно продольной оси нити. Менее чем через 1 мс после этого под влиянием актомиозинового комплекса происходит гидролиз АТФ и отщепление его продуктов (АДФ и неорганического фосфата). При этом угол наклона мостика относительно продольной оси нити изменяется до 40°. Такой конформационный переход, происходящий в области прикрепления головки миозиновой молекулы, обусловливает развитие усилия и смещение тонких филаментов к центру саркомера. Размыкание мостика. Связывание новой молекулы АТФ с мостиком вызывает его отделение от тонкого филамента. Мостик размыкается, возвращаясь в прежнее положение относительно миозиновой нити и может прийти в замыкание со следующим активным центром на тонкой. Вопрос 36. Поперечнополосатая сердечная мышечная ткань. Структурно-функциональная характеристика кардиомиоцитов. Сердечная мышечная ткань образована клетками - кардиомиоцитами, связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон. В отличии от скелетной, она представлена не симпластом, а синцитием. Кардиомиоциты - цилиндрические или ветвящиеся клетки, более крупные в желудочках. Кардиомиоциты содержат одно или два (в основном 2) ядра и саркоплазму, покрыта сарколеммой, которая снаружи окружена базальной мембраной. Ядра кардиомиоцитов - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок. Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов: многочисленные интердигитации, десмосомы, полоски слипания, связь за счёт гликокаликса. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения, обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения. Кардиомиоциты бывают 5 видов: Рабочие (сократительные) – образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим большую часть их саркоплазмы. Синусные (пейсмейкерные) – за счёт них происходит создание импульса. Они занимают центральную часть синусного узла. В их цитоплазме наблюдается незначительное количество миофибрилл. Проводящие – проводят импульс, образуя цепочки клеток, соединенных своими концами. Переходные – служат промежуточным звеном между синусными и проводящими и между проводящими и рабочими. Секреторные – вырабатывают натрийуретический фактор (гормон), участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Вопрос 37. Общая характеристика гладкой мышечной ткани. Типы гладких миоцитов. Строение, локализация и функции. Гладкие мышцы выстилают некоторые внутренние органы, такие как кишечник, матка, мочевой пузырь, а также сосуды. Кроме того, из них формируются сфинктеры и связки. Структурной единицей гладкой мышечной ткани являются гладкие миоциты (ГМК), сократительный аппарат которых представлен не миофибриллами, а протофибриллами, которые также построены из актина и миозина. Структурной единицей протофибриллы является также саркомер. По происхождению ГМК делятся на мезенхимные, эпидермальные и нейральные: Мышечная ткань мезенхимного происхождения. Гладкий миоцит – веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены около полюсов ядра. Концы протофиламентов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Гладкие миоциты располагаются без заметных межклеточных пространств. На концах клеток плазмолемма образует узкие трубчатые впячивания. Миоциты разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Мышечная ткань эпидермального происхождения. Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки тоже восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках – сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа Мышечная ткань нейрального происхождения. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок. Вопрос 38. Морфофункциональная характеристика гладкого миоцита (СМ и ЭМ). Сократительный и опорный аппарат. Механизм мышечного сокращения. Гладкие миоциты - одноядерные клетки преимущественно веретеновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом. Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной, содержат одно ядро и саркоплазму, в которой располагаются органеллы и включения. Ядро гладких миоцитов - сигарообразной формы, расположено вдоль длинной оси клетки в ее центральной утолщенной части. Ядро обычно диплоидное, в нем преобладает эухроматин, выявляются 1-2 ядрышка. Саркоплазма гладких миоцитов содержит умеренно развитые органеллы общего значения, которые располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята миофиламентами. В саркоплазме выделяют следующие аппараты: Сократительный Передачи возбуждения (с сарколеммы на сократительный аппарат) Опорный Энергетический Синтетический Лизосомальный Сокращение гладких миоцитов обеспечивается взаимодействием актиновых и миозиновых миофиламентов и развивается в соответствии с моделью скользящих нитей. Оно происходит медленнее и длится дольше, чем в скелетной мышце, что обусловлено более низкой скоростью гидролиза АТФ в гладких миоцитах. Сократительный аппарат гладких миоцитов представлен тонкими (актиновыми) и толстыми (миозиновыми) филаментами, которые, однако, в отличие от поперечнополосатых мышечных тканей, не формируют миофибрилл. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са2+ в саркоплазму, который в этих клетках выделяется саркоплазматической сетью и кавеолами (колбовидные впячивания поверхности сарколеммы, содержащие высокие концентрации кальция), а также вследствие увеличения проницаемости сарколеммы для данных ионов. Опорный аппарат гладкого миоцита представлен его сарколеммой, базальной мембраной, системой элементов цитоскелета и связанных с ними плотных телец. Плотные тельца – овальные или веретеновидные структуры, лежащие вдоль длинной оси миоцита свободно в его саркоплазме или связанные с внутренней поверхностью сарколеммы. Механизм мышечного сокращения такой же, как в вопросе 35. Вопрос 39. Нервная ткань. Гистогенез. Производные нервной трубки (нейробласты, глиобласты), нервного гребня и нейральных плакод. Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой. Гистогенез. Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней эпидермальной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных или нейроэпителиальных клеток. Вентрикулярные клетки являются предшественниками нейронов и клеток макроглии Далее клетки дифференцируются и образуются субвентрикулярная и промежуточная зоны. Промежуточная зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем превращаются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Маргинальная зона (краевая вуаль) формируется из врастающих в нее аксонов нейробластов и глии и дает начало белому веществу. | ||||||||||||||||||