Общая. Эпителиальные ткани
 Скачать 4.65 Mb. Скачать 4.65 Mb.
|

|
Классификация кроветворных клеток: Плюрипотентные стволовые клетки крови по строению напоминают малые лимфоциты, но могут быть идентифицированы иммуноцитохимически по набору антигенов на клеточной поверхности. СКК сосредоточены у взрослого человека преимущественно в красном костном мозге, однако обнаруживаются в крови, циркулируя в которой они попадают в другие органы кроветворения. Частично детерминированные поли-, или мультипотентные родоначальные клетки, именуемые также полустволовыми клетками. Развиваются из СКК, способны к ограниченному самоподдержанию. Родоначальные клетки, относящиеся к данному и следующему (3) классам, называют также колониеобразующими единицами (КОЕ). Частично детерминированные полипотентные родоначальные клетки включают родоначальную клетку лимфоцитоноэза (KOE-JI) и родоначальную клетку миелопоэза (КОЕ-ГЭММ), дающую начало гpaнулоцитам, эритроцитам, моноцитам и мегакариоцитам. Унипотентные родоначальные клетки, прошедшие новый этап коммитирования и детерминированные в направлении развития только одного вида форменных элементов (за исключением КОЕ-ГМ, дающий КОЕ-Г(Н), т.е. родоначальную клетку гранулоцитов (нейтрофильных) и КОЕ-Мо – родоначальную клетку моноцитов). Морфологически распознаваемые предшественники – бластные формы. Представляют отдельные линии развития форменных элементов. Пролиферативная активность этих клеток ограничена; способностью к самоподдержанию они не обладают. Созревающие (дифференцирующиеся) клетки. Подвергаются структурной и функциональной дифференцировке, образуя соответствующий вид форменных элементов, в ходе которой они (за исключением лимфоцитов и моноцитов) утрачивают способность к делению. Зрелые (дифференцированные) форменные элементы, циркулирующие в крови. Неспособны к делению (за исключением лимфоцитов и моноцитов) СКК по строению напоминают малые лимфоциты, но могут быть идентифицированы иммуноцитохимически по набору антигенов на клеточной поверхности. Наиболее типичным маркерным признаком служит сочетание CD34+CD38. Локализация СКК. СКК сосредоточены у взрослого человека преимущественно в красном костном мозге, однако обнаруживаются в крови, циркулируя в которой они попадают в другие органы кроветворения. Очевидно, что СКК обладают уникальной для малодифференцированных клеток способностью выселяться из костного мола и, подобно зрелым форменным элементам, мигрировать через эндотелий венозных синусов в кровь. В красном костном мозге их содержание невелико (одна СКК приходится примерно на 2000 клеток), в циркулирующей крови СКК составляют 0.0001% от общего числа лейкоцитов. Основные свойства: Обладают способностью к самоподдержанию без притока клеток извне, т.е. к образованию в результате деления дочерних клеток, не отличающихся своим практически неограниченным пролиферативным потенциалом от родительской. Редко делятся (основное состояние - покоя); однако могут быть вовлечены в пролиферацию при значительных кровопотерях и при воздействии факторов роста. Способны образовывать все виды форменных элементов крови. Устойчивы к действию повреждающих факторов Располагаются в местах, хорошо защищенных от внешних воздействий (ячейки в костной ткани) и обладающих обильным кровоснабжением Циркулируют в крови, мигрируя в другие органы кроветворения Вопрос 15. Красный костный мозг. Строение. Эритроцитопоэз. Основные стадии развития и дифференцировки эритроцитов. Красный костный мозг является кроветворной частью костного мозга. Он заполняет губчатое вещество плоских и трубчатых костей. Стромой костного мозга является ретикулярная ткань, образующая микроокружение для кроветворных клеток. В настоящее время к элементам микроокружения относят также остеогенные, жировые, адвентициальные, эндотелиальные клетки и макрофаги. Ретикулярные клетки благодаря своей отростчатой форме выполняют механическую функцию, секретируют компоненты основного вещества – преколлаген, гликозаминогликаны, проэластин и микрофибриллярный белок и участвуют в создании кроветворного микроокружения, специфического для определенных направлений развивающихся гемопоэтических клеток, выделяя ростовые факторы. Остеогенными клетками называют стволовые клетки опорных тканей, остеобласты и их предшественники. Остеогенные клетки также способны вырабатывать ростовые факторы, индуцировать родоначальные гемопоэтические клетки в местах своего расположения к пролиферации и дифференцировке. Адипоциты – жировые клетки, являются постоянными элементами костного мозга. Адвентициальные клетки сопровождают кровеносные сосуды и покрывают более 50 % наружной поверхности синусоидных капилляров. Под влиянием гемопоэтинов и др. факторов они способны сокращаться, что способствует миграции клеток в кровоток. Эндотелиальные клетки сосудов костного мозга принимают участие в организации стромы и процессов кроветворения, синтезируют коллаген IV типа, гемопоэтины. Эндотелиоциты способны к сократительным движениям, которые способствуют выталкиванию клеток крови в синусоидные капилляры. Эндотелиоциты выделяют колониестимулирующие факторы (КСФ) и белок с антигенными свойствами — фибронектин, обеспечивающий прилипание клеток друг к другу и субстрату. Макрофаги в костном мозге представлены неоднородными по структуре и функциональным свойствам клетками. Некоторые из популяций макрофагов секретируют ряд биологически активных веществ (эритропоэтин, колониестимулирующий фактор, интерлейкины, простагландины, интерферон и др.). Макрофаги при помощи своих отростков, проникающих через стенки синусов, улавливают из кровотока железосодержащие соединения и далее передают его развивающимся эритроидным клеткам для построения гема. Межклеточное вещество. В костном мозге это вещество содержит коллаген II, III и IV типа, гликопротеины, протеогликаны и др. Эритроцитопоэз – процесс образования и созревания эритроцитов, происходящий в миелоидной ткани. Ход развития эритроцитов из стволовой клетки крови описывается последовательностью: Стволовая клетка крови (СКК) Колониеобразующая единица (КОЕ-ГЭММ) КОЕ-Э. 1, 2 и 3 морфологически не различаются, отличаются только пролиферативной активностью и чувствительностью к факторам эртроцитопоэза. Проэритробласт - крупная клетка (диаметром около 18-22 мкм) с большим сферическим ядром, содержащим мелкодисперсный хроматин и 2-3 бледных ядрышка. Клетка интенсивно пролиферирует, давая начало эритробластам. Базофилъный эритробласт - меньших размеров, чем проэритробласт, с более мелким ядром, содержащим умеренно конденсированный хроматин и ядрышки. Цитоплазма резко базофильна благодаря высокому содержанию полисом, активно синтезирующих гемоглобин. Базофильные эритробласты активно делятся. Полихроматофильный эритробласт характеризуется более мелкими размерами, чем базофильный эритробласт, его ядро более компактно, чем в базофильном эритробласте, глыбки хроматина в нем распределены в виде спиц колеса, ядрышко не выявляется. Цитоплазма окрашивается полихромно: она воспринимает как основные красители (вследствие наличия в ней многочисленных полисом), так и кислые (из-за накопления в ней оксифилыю окрашивающегося гемоглобина. Оксифилъный эритробласт образуется путем дифференцировки из полихроматофильного эритробласта. По размерам он чуть крупнее эритроцита. Обладает оксифильно окрашенной цитоплазмой, богатой гемоглобином, в которой органеллы почти полностью отсутствуют. Ядро мелкое, компактное, расположено эксцентрически. Способность к делению теряется. Выталкиванию ядра предшествует перестройка цитоскелета клетки, образующего структуру в виде манжетки, которая способствует активному выталкиванию ядра с тонким ободком окружающей его цитоплазмы за пределы клетки, где оно быстро фагоцитируется макрофагами. Ретикулоцит представляет собой безъядерную (постклеточную) структуру. Его оксифильная цитоплазма, почти целиком заполненная гемоглобином, содержит остатки полирибосом и других органелл. Эритроцит превщарается из ретикулоцита в кровотоке примерно через 1-2 дня после нахождения в кровотоке. Вопрос 16. Гранулоцитопоэз. Стадии развития и дифференцировки гранулоцитов. Гранулоцитопоэз - образование и дифференцировка гранулоцитов - происходит в ККМ. Исходным источником развития всех гранулоцитов служит СКК, которая дает начало КОЕ-ГЭММ. Последовательность начальных этапов развития гранулоцитов: Нейтрофильных: СКК -> КОЕ-ГЭММ -> КОЕ-ГМ -> КОЕ-Г(Н) Базофильных: СКК -> КОЕ-ГЭММ -> КОЕ-Баз Эозинофильных: СКК -> КОЕ-ГЭММ -> КОЕ-Эо. Последующие стадии развития гранулоцитов протекают для всех трех типов клеток однотипно: миелобласт -> промиелоцит -> миелоцит -> метамиелоцит (юный гранулоцит) -> палочкоядерный гранулоцит -> сегментоядерный гранулоцит. Процесс дифференцировки предшественников гранулоцитов в зрелые клетки включает: Уменьшение размеров клетки Снижение, а в дальнейшем (со стадии метамиелоцита) - утрату способности к делению Изменение формы ядра - от округлой до бобовидной и палочковидной, его сегментация; нарастание конденсации ядерного хроматина Выработку и накопление гранул в цитоплазме Нарастание подвижности клетки, обусловленное перестройкой цитоскелета Приобретение разнообразных рецепторов плазмолеммы Стадии дифференцировки: Миелобласт – крупная (диаметр - 16-24 мкм) клетка с развитой слабобазофильной цитоплазмой и большим круглым светлым ядром. Имеет хорошо развитый синтетический аппарат, необходимый для синтеза гранул. Промиелоцит – клетка морфологически схожая с миелобластом. Промиелоцит делится митотически. К концу этой стадии образование азурофильных гранул завершается. Поэтому, поскольку промиелоцит активно делится, на последующих стадиях содержание этих гранул непрерывно снижается. Миелоцит обычно меньших размеров, чем промиелопит (диаметр - 10-16 мкм). Характеризуется небольшой инвагинацией ядра с более крупными гранулами гетерохроматина; ядрышки исчезают. Цитоплазма содержит первичные гранулы (количество которых постепенно падает) и вторичные (специфические) гранулы (число которых непрерывно возрастает благодаря деятельности синтезирующих их грЭПС и КГ). К концу стадии миелоцита вторичных гранул становится больше, чем первичных. Миелоцит - последняя митотически активная клетка линии гранулоцитов. Метамиелоцит - меньших размеров (по сравнению с миелоцитом) и отличается от него более заметной инвагинацией ядра, которое принимает бобовидную форму и уплотняется. По ходу созревания метамиелоцита, а позднее на стадиях палочко- и сегментоядерного гранулоцита происходят дальнейшие изменения ядра и цитоплазмы Ядро изменяет форму - из бобовидного становится подковообразным, а затем палочковидным с последующим формированием перетяжек, разделяющих его на сегменты; конденсация хроматина прогрессивно нарастает. Цитоплазма характеризуется уменьшением количества элементов грЭПС, рибосом, митохондрий, редукцией комплекса Гольджи, отмечаются количественные и качественные изменения цитоскелета (обеспечивающие высокую подвижность, образование псевдоподий) и рецепторного аппарата плазмолеммьг (обусловливающие высокую способность к хемотаксису, фагоцитозу, адгезивным взаимодействиям и распознаванию различных сигнальных молекул) Вопрос 17. Красный костный мозг. Строение. Стадии развития тромбоцитов. ККМ и его строение в вопросе 15. Тромбоцнтопоэз - процесс образования и созревания тромбоцитов, происходящий в миелоидной ткани. Тромбоциты образуются в результате процесса частичной фрагментации цитоплазмы гигантских клеток костного мозга - мегакариоцитов. Ход развития мегакариоцитов из стволовой клетки крови описывается последовательностью: СКК -> КОЕ-ГЭММ -> КОЕ-Мег -> мегакариобласт -> промегакариоцит -> мегакариоцит -> тромбоцит. СКК, КОЕ-ГЭММ, КОЕ-Мег – это активно делящиеся клетки, морфологически схожи друг с другом, отличающиеся в основном активностью пролиферации и ответом на различные стимулирующие факторы. Мегакариобласт – клетка диаметром 15-25 мкм, имеет ядро с инвагинациями и относительно небольшой ободок базофильной цитоплазмы. Клетка способна к делению митозом, иногда содержит два ядра. При дальнейшей дифференцировке утрачивает способность к митозу и делится путем эндомитоза, при этом увеличиваются плоидность и размер ядра. Промегакариоцит – клетка диаметром 30-40 мкм, содержит тетраплоидные, октоплоидные ядра (4п, 8п), несколько пар центриолей. Объем цитоплазмы возрастает, в ней начинают накапливаться азурофильные гранулы. Клетка также способна к эндомитозу и дальнейшему увеличению полиплоидности ядер. Мегакариоцит – дифференцированная форма. Среди них различают резервные клетки, не образующие пластинок, и зрелые активированные клетки, образующие кровяные пластинки. Мегакариоцит при созревании из мегакариобласта становится крупнее, достигая 50-100 мкм в диаметре; его ядро и цитоплазма претерпевают выраженные изменения. Дифференцировка ядра включает активную репликацию ДНК без митоза. Полиплоидные клетки в дальнейшем претерпевают эндомитоз с образованием многочисленных связанных перемычками долей ядра, в котором общее содержание ДНК соответствует 4-128n (наиболее часто - 16 или 32n). Хроматин постепенно конденсируется, ядрышко, как правило, не выявляется. Размеры клетки обычно соответствуют степени ее полиплоидизации. Дифференцировка цитоплазмы мегакариоцитов начинается только по завершении репликации ДНК. Наиболее заметными ее проявлениями служат: Разделение цитоплазмы на три зоны: Околоядерная зона содержит элементы грЭПС, хорошо развитый комплекс Гольджи, митохондрии и центриоли. Промежуточная зона - наибольшая по ширине, содержит гранулы и систему мембранных демаркационных каналов – каналов, разрезающих цитоплазму мегакариоцитов на территории размером 2-4 мкм, соответствующие границам будущих тромбоцитов и содержащие гранулы. Краевая (периферическая) зона свободна от большинства органелл и гранул, в ней в значительном количестве сосредоточены элементы цитоскелета. Образование и накопление гранул, характерных для тромбоцитов и содержащих типичные для них белки Образование филоподий (протромбоцитов) – узких длинных лентовидных отростков мегакариоцитов, которые через поры эндотелия синусов красного костного мозга проникают в их просвет, где распадаются на отдельные тромбоциты. Вопрос 18. Моноцитопоэз. Основные стадии развития и дифференцировки моноцитов. Моноцнтопоэз - процесс развития моноцитов - происходит в красном костном мозге и описывается последовательностью: СКК -> КОЕ-ГЭММ -> КОЕ-ГМ -> КОЕ-М -> монобласт -> промонопит -> моноцит СКК, КОЕ-ГЭММ, КОЕ-ГМ, КОЕ-М – это активно делящиеся клетки, морфологически схожи друг с другом, отличающиеся в основном активностью пролиферации и ответом на различные стимулирующие факторы. Промоноцит - сравнительно крупная клетка (диаметром 12-18 мкм) с большим светлым слегка вогнутым ядром, в котором располагаются 1-2 ядрышка. Базофильная цитоплазма содержит умеренно развитую грЭПС, полисомы, митохондрии, центриоли и крупный комплекс Гольджи, от которого отделяются незрелые азурофильные гранулы. Промоноциты делятся и постепенно дифференцируются в моноциты. Процесс преобразования монобластов в моноциты включает: Дальнейшее увеличение размеров клетки преимущественно за счет нарастания объема цитоплазмы Снижение базофилии цитоплазмы Накопление в ней азурофильных гранул (лизосом) Изменение формы ядра, которое становится бобовидным Выделяясь в синусы ККМ, они попадают в кровь, в которой циркулируют от 8 часов до 3-4 суток, а далее через стенку сосудов мигрируют в ткани. Лишь около 5% моноцитов, имеющихся в организме, циркулирует в крови, остальные находятся во внесосудистом пуле. В тканях они превращаются в различные виды макрофагов (вместе с которыми образуют единую моноцитарно-макрофагальную систему), а также в дендритные антиген-представляющие клетки Вопрос 19. Т-лимфоциты: субпопуляции. Характеристика рецепторов. Антигеннезависимая и антигензависимая пролиферация и дифференцировка. Т-лимфоциты — самая многочисленная популяция лимфоцитов, составляющая 70—90 % лимфоцитов крови. Эти клетки имеют на своей плазмолемме маркеры (CD3,4,8), позволяющие идентифицировать их, а также ТКР (Т-клеточные рецепторы) – специфические антигенраспознающие рецепторы. Субпопуляции Т-лимфоцитов: Т-хелперы – стимулируют дифференцировку В-лимфоцитов, образование из них плазмоцитов и продукцию иммуноглобулинов. CD3 + CD4 + ТКР Т-супрессоры – способны ингибировать действие Т-хелперов, В-лимфоцитов, плазмоцитов и регулировать силу иммунного ответа. CD3 + CD8 + ТКР Т-киллеры – участвуют в реакциях клеточного иммунитета, обеспечивая разрушение (лизис) чужеродных клеток и собственных измененных клеток (например, опухолевых клеток). CD3 + CD8 + ТКР Т-памяти – долгоживущие клетки, которые активируются при повторном попадании чужеродного объекта в организм. CD45RO+ и ТКР. Клетка памяти на своей плазмолемме имеет много ТКР, потому что должна активироваться даже при маленьких концентрациях антигена. 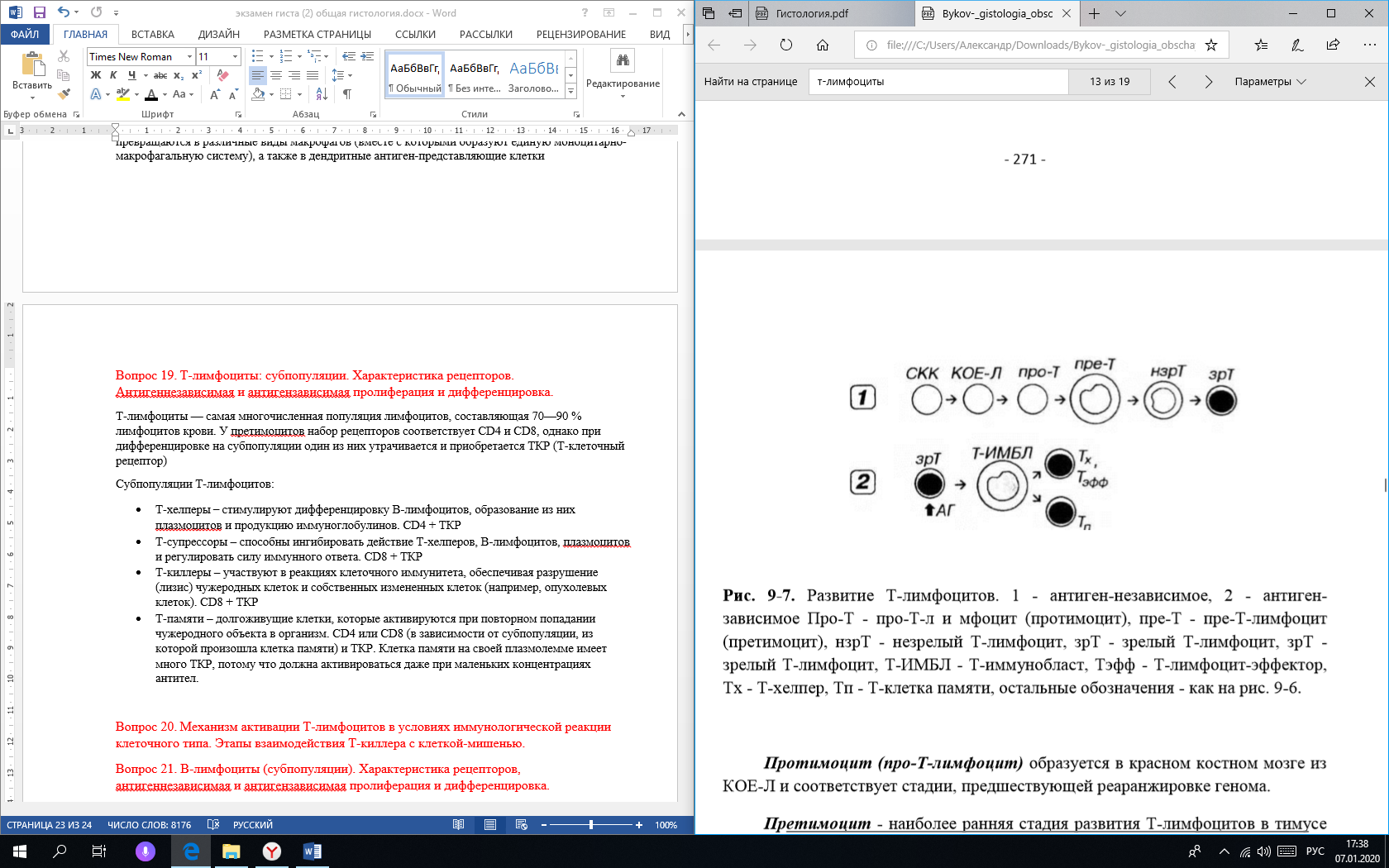 Антигеннезависимая пролиферация. |