Ферменты и коферменты, заполнив таблицу
 Скачать 0.74 Mb. Скачать 0.74 Mb.
|

|
тема: Распад и синтез олеиновой кислоты Ответ составьте по следующему плану:
Содержится во многих животных жирах в виде сложных эфиров — глицеридов: говяжьем жире (41—42 %), в свином (37—44 %), в тресковом (30 %), а также содержится во многих растительных маслах: масло сасанквы (85—89 %), масло лесного ореха (70—84 %), масло камелии (80 %), масло бурити (Mauritia Vinifera) (79 %), масло папайи (79 %), масло марулы (70—78 %), масло моринга (71 %), миндальное масло (64—82 %), оливковое масло (55,0—83,0 %), масло асаи (60 %), масло орехов Кешью (60 %), масло авокадо (59—75 %), масло абрикосовых косточек (58—74 %), арахисовое масло (66 %), масло Персиковых косточек (55—75 %), фисташковое масло (51—54 %), масло орехов макадамии (50—67 %), масло мякоти пекуи (54 %), масло андиробы (Carapa guianensis) (50,5 %), масло дерева ним (50 %), масло путерии (50 %), Аргановое масло (48 %), масло косточек пекуи (46 %), овсяное масло (46 %), масло калодендрума капского (Calodendrum capense) (45 %), масло понгамии (44,5—71 %), масло орехов пекан (43—51 %), масло купуасу (44 %), масло манго (43 %), масло авелланского ореха (42 %), масло ши (40—45 %), подсолнечное масло (14,0—39,4 %, высокоолеиновое 61,0—69,8 %), горчичное масло (22—30 %), масло бразильского ореха (38 %), масло дерева сал (37—43 %), кунжутное масло (37—42 %), тыквенное масло (35—47 %), пальмовое масло (35—45 %), масло какао (34—36 %), масло расторопши (30 %), кукурузное масло (24—42 %), масло кокум (Garcinia indica) (30—42 %), масло орехов иллипа (Shorea stenoptera) (32—38 %), рисовое масло (32—38 %).
Переваривание жиров в кишечнике. Небольшое количество триглицеридов переваривается в желудке под действием лингвальной липазы, которая секретируется железами языка в ротовой полости и проглатывается вместе со слюной. Количество перевариваемых таким образом жиров составляет менее 10%, а потому не существенно. Основное переваривание жиров происходит в тонком кишечнике, о чем сказано далее. Эмульгирование жиров желчными кислотами и лецитином. Первый этап переваривания жиров заключается в физическом разрушении капель жира на мелкие частицы, поскольку водорастворимые ферменты могут действовать только на поверхности капли. Этот процесс называют эмульгированием жиров, он начинается в желудке с перемешивания жиров с другими продуктами переваривания желудочного содержимого. Далее основной этап эмульгирования происходит в двенадцатиперстной кишке под влиянием желчи, секрета печени, который не содержит пищеварительных ферментов. Однако желчь содержит большое количество желчных солей, а также фосфолипид -- лецитин. Эти компоненты, в особенности лецитин, чрезвычайно важны для эмульгирования жиров. Полярные частицы (места, в которых происходит ионизация воды) желчных солей и молекул лецитина хорошо растворимы в воде, тогда как большая оставшаяся часть этих молекул хорошо растворима в жирах. Таким образом, жирорастворимые порции секрета печени растворяются в поверхностном слое жировых капель вместе с выступающей полярной частью. В свою очередь, выступающая полярная часть растворима в окружающей водной фазе, что значительно снижает поверхностное натяжение жиров и делает их также растворимыми. Когда поверхностное натяжение капли нерастворимой жидкости низкое, нерастворимая в воде жидкость во время перемещения значительно легче разрушается на множество мелких частиц, чем при более высоком поверхностном натяжении. Следовательно, основная функция желчных солей и лецитина -- делать капли жира способными к легкому размельчению при перемешивании с водой в тонком кишечнике. Это действие аналогично действию синтетических моющих средств, широко используемых в домашнем хозяйстве для устранения жира. Каждый раз в результате перемешивания в тонком кишечнике диаметр жировых капель существенно уменьшается, поэтому общая жировая поверхность многократно увеличивается. Из-за того, что средний диаметр частичек жира в кишечнике после эмульгирования оказывается менее 1 мкм, общая площадь жировой поверхности, образованная в результате процесса эмульгирования, увеличивается в 1000 раз. Фермент липаза является водорастворимым и может воздействовать только на поверхности жировых капель. Отсюда понятно, насколько значительна детергентная роль лецитина и желчных солей в переваривании жиров. В процессах пищеварения все омыляемые липиды (жиры, фосфолипиды, гликолипиды, стериды) подвергаются гидролизу на составные части, уже названные ранее, стерины же химическим изменениям не подвергаются. При изучении этого материала следует обратить внимание на отличия пищеварения липидов от соответствующих процессов для углеводов и белков: особую роль желчных кислот в распаде липидов и транспорте продуктов пищеварения. В составе липидов пищи преобладают триглицериды. Фосфолипидов, стреинов и других липидов потребляется значительно меньше. Большая часть поступающих с пищей триглицеридов расщепляется до моноглицеридов и жирных кислот в тонком кишечнике. Гидролиз жиров происходит под влиянием липаз сока поджелудочной железы и слизистой оболочки тонкого кишечника. Соли желчных кислот и фосфолипиды, проникающие из печени в просвет тонкого кишечника в составе желчи, способствуют образованию устойчивых эмульсий. В результате эмульгирования резко увеличивается площадь соприкосновения образовавшихся мельчайших капелек жира с водным раствором липазы, и этим самым увеличивается липолитическое действие фермента. Соли желчных кислот стимулируют процесс расщепления жиров не только участвуя в их эмульгировании, но и активируя липазу. Расщепление стероидов происходит в кишечнике при участии фермента холинэстеразы, выделяющегося с соком поджелудочной железы. В результате гидролиза стероидов образуются жирные кислоты и холестерин. Фосфолипиды расщепляются полностью или частично под действием гидролитических ферментов - специфических фосфолипаз. Продуктом полного гидролиза фосфолипидов являются: глицерин, высшие жирные кислоты, фосфорная кислота и азотистые основания. Всасыванию продуктов переваривания жиров предшествует образование мицелл - надмолекулярных образований или ассоциатов. Мицеллы содержат в качестве основного компонента соли желчных кислот, в которых растворены жирные кислоты, моноглицериды, холестерин и т.п. В клетках кишечной стенки из продуктов пищеварения, а в клетках печени, жировой ткани и других органов из предшественников, возникших в обмене углеводов и белков, происходит построение молекул специфических липидов тела человека - ресинтез триглицеридов и фосфолипидов. Однако их жирнокислотный состав по сравнению с жирами пищи изменен: в триглицеридах, синтезируемых в слизистой оболочке кишечника содержатся арахидоновая и линоленовая кислоты даже в том случае, если они отсутствуют в пище. Кроме того, в клетках кишечного эпителия жировая капля покрывается белковой оболочкой и происходит формирование хиломикронов - большая жировая капля, окруженная небольшим количеством белка. Транспортирует экзогенные липиды в печень, адипозную ткань, соединительную ткань, в миокард. Поскольку липиды и некоторые их составные части нерастворимы в воде, для переноса из одного органа в другой они образуют особые транспортные частицы, в составе которых обязательно есть белковый компонент. В зависимости от места образования эти частицы различаются структурой, соотношением составных частей и плотностью. Если в составе такой частицы в процентном соотношении жиры преобладают над белками, то такие частицы называются липопротеинами очень низкой плотности (ЛПОНП) или липопротеинами низкой плотности (ЛПНП). По мере увеличения процентного содержания белка (до 40%) частица превращается в липопротеин высокой плотности (ЛПВП). В настоящее время изучение таких транспортных частиц дает возможность с большой степенью точности оценивать состояние липидного обмена организма и использование липидов в качестве источников энергии. Если образование липидов происходит из углеводов или белков, предшественником глицерина становится промежуточный продукт гликолиза - фосфодиоксиацетон, жирных кислот и холестерина - ацетилкофермент А, аминоспиртов - некоторые аминокислоты. Синтез липидов требует больших энерготрат для активации исходных веществ. Основной частью продуктов распада жиров всасывается из клеток кишечного эпителия в лимфатическую систему кишечника, грудной лимфатический проток и только затем - в кровь. Незначительная часть короткоцепочечных жирных кислот и глицерина способна всасываться непосредственно в кровь воротной вены. Липиды, образовавшиеся из продуктов пищеварения, поступают, в основном, в депо, где откладываются в запас. Они могут мобилизоваться при увеличении потребности организма в них. Часть вновь синтезированных липидов поступает в клетки различных органов, где используется преимущественно как структурный компонент протоплазмы и мембран клеток. Эти липиды, в отличие от депонированных, обладают видовой специфичностью и значительной устойчивостью. Мобилизация липидов из депо особенно усиливается при охлаждении организма, длительной мышечной работе, понижении содержания углеводов. Мобилизация представляет собою липолиз (гидролитическое расщепление) липидов и включение продуктов этого расщепления в обменные процессы в различных органах. 2. Всасывание жиров Переваривание жиров в желудочно-кишечном тракте (ЖКТ) отличается от переваривания белков и углеводов. Жиры не растворимы в жидкой среде кишечника, и поэтому для того, чтобы они гидролизовались и всасывались, необходимо их эмульгирование -- разбивка на мельчайшие капельки. В результате получается эмульсия - дисперсия микроскопических частиц одной жидкости в другой. Эмульсии могут быть образованы двумя любыми не смешивающимися жидкостями. В большинстве случаев одной из фаз эмульсий является вода. Эмульгирование жиров идёт с помощью желчных кислот, которые синтезируются из холестерина в печени. Так что холестерин важен для усвоения жиров. Как только произошло эмульгирование, жиры (липиды) становятся доступными для панкреатических липаз, которые секретирует поджелудочная железа, особенно для липазы и фосфолипазы А2. Продукты расщепления жиров панкреатическими липазами -- это глицерин и жирные кислоты. В результате расщепления молекул липидов (жиров) получаются глицерин и жирные кислоты. Они, а также мельчайшие капли нерасщеплённого эмульгированного жира, всасываются в верхнем отделе тонкого кишечника в начальных 100 см. В норме всасывается 98% пищевых липидов. 1. Короткие жирные кислоты (не более 10 атомов углерода) всасываются и переходят в кровь без каких-либо особенных механизмов. Этот процесс важен для грудных детей, т.к. молоко содержит в основном коротко- и среднецепочечные жирные кислоты. Глицерол тоже всасывается напрямую. 2. Другие продукты переваривания (жирные кислоты, холестерол, моноацилглицеролы) образуют с желчными кислотами мицеллы с гидрофильной поверхностью и гидрофобным ядром. Их размеры в 100 раз меньше самых мелких эмульгированных жировых капелек. Через водную фазу мицеллы мигрируют к щеточной каемке слизистой оболочки. Здесь мицеллы распадаются и липидные компоненты проникают внутрь клетки, после чего транспортируются в эндоплазматический ретикулум. Желчные кислоты частично также могут попадать в клетки и далее в кровь воротной вены, однако большая их часть остается в химусе и достигает подвздошной кишки, где всасывается при помощи активного транспорта. 3.Этапы переваривания жиров Потребность в липидах взрослого организма составляет 80-100 г в сутки, из них растительных (жидких) жиров должно быть не менее 30%. С пищей в основном поступают триацилглицеролы, фосфолипиды и эфиры ХС. Переваривание липидов осложняется тем, что их молекулы полностью или частично гидрофобны. Для преодоления этой помехи используется процесс эмульгирования, когда гидрофобные молекулы (ТАГ, эфиры ХС) или гидрофобные части молекул (ФЛ, ХС) погружаются внутрь мицеллы, а гидрофильные остаются на поверхности, обращенной к водной фазе. Условно внешний обмен липидов можно подразделить на следующие этапы: 1. Эмульгирование жиров пищи - необходимо для того, чтобы ферменты ЖКТ смогли начать работу. 2. Гидролиз триацилглицеролов, фосфолипидов и эфиров ХС под влиянием ферментов ЖКТ. 3. Образование мицелл из продуктов переваривания (жирных кислот, МАГ, холестерола). 4. Всасывание образованных мицелл в эпителий кишечника. 5. Ресинтез триацилглицеролов, фосфолипидов и эфиров ХС в энтероцитах. После ресинтеза липидов в кишечнике они собираются в транспортные формы - хиломикроны (основные) и липопротеины высокой плотности (ЛПВП) (малое количество) - и разносятся по организму.
Катаболизм идет в три этапа:
Анаболизм: синтез триглицеридов.
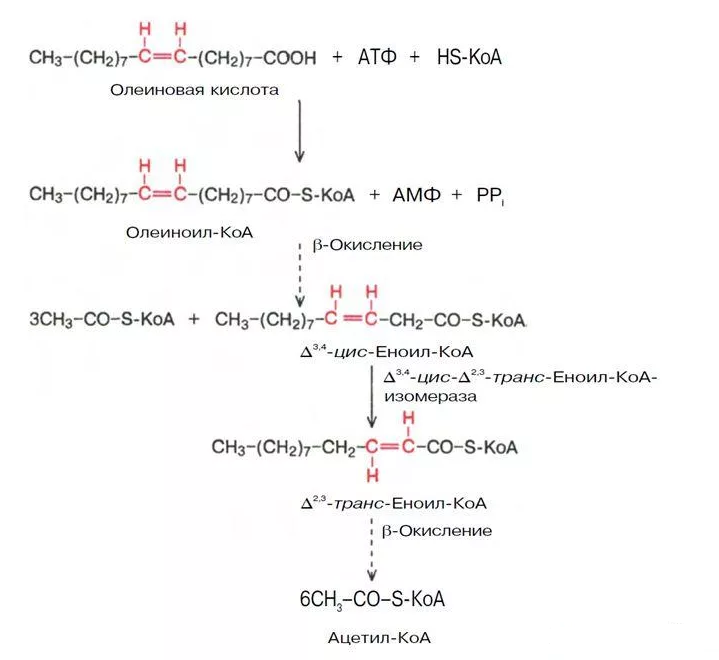 Так как число атомов углерода равно 18, то количество молекул ацетил-S-КоА равно 9. Значит при его окислении в ЦТК образуется 9×12=108 молекул АТФ. исходя из формулы (n/2 - 1) число циклов β-окисления равно 8. При расчете получаем 8×5=40 молекул АТФ. в кислоте имеются 1 двойная связи. Следовательно, в двух циклах β-окисления не образуется 1 молекула ФАДН2, что равноценно потере 2 молекул АТФ. на активацию жирной кислоты тратятся 2 макроэргические связи. таким образом, энергетический выход 108 + 40 - 2 - 2 =144 молекулы АТФ.
На первой стадии из двух молекул ацетил-КоА образуется ацетоацетил-КоА: Реакция катализируется ферментом ацетил-КоА-ацетилтрансферазой (ацетоацетил-КоА-тиолазой). Затем ацетоацетил-КоА взаимодействует еще с одной молекулой ацетил-КоА. Реакция протекает под влиянием фермента гидроксиметилглутарил-КоА-синтазы: 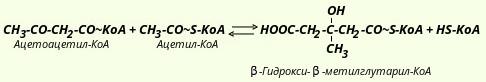 Образовавшийся β-гидрокси-β-метилглутарил-КоА способен под действием гидроксиметилглутарил-КоА-лиазы расщепляться на ацетоацетат и ацетил-КоА: 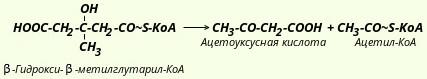 Ацетоуксусная кислота способна восстанавливаться при участии НАД-зависимой D-β-гидроксибутиратдегидрогеназы; при этом образуется D-β-гидроксимасляная кислота (D-β-гидроксибутират). Следует еще раз подчеркнуть, что фермент специфичен по отношению к D-стереоизомеру и не действует на КоА-эфиры: Существует также и второй путь синтеза кетоновых тел. Образовавшийся путем конденсации двух молекул ацетил-КоА ацетоацетил-КоА способен отщеплять коэнзим А и превращаться в свободную ацетоуксусную кислоту. Этот процесс катализируется ферментом ацетоацетил-КоА-гидролазой (деацилазой): Однако второй путь образования ацетоуксусной кислоты не имеет существенного значения, так как активность деацилазы в печени низкая. В крови здорового человека кетоновые тела содержатся лишь в очень небольших концентрациях. При патологических состояниях (например, у лиц с тяжелой формой сахарного диабета, при голодании, а также у животных с острым аллоксановым диабетом) концентрация кетоновых тел в крови увеличивается и может достигать 20 ммоль. Такое состояние, которое получило название кетоза, возникает в тех случаях, когда скорость образования кетоновых тел превышает способность периферических тканей их утилизировать. В последние годы начали постепенно накапливаться данные, указывающие на важную роль кетоновых тел в поддержании энергетического гомеостаза. Кетоновые тела - своего рода поставщики топлива для мышц, мозга, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала. В периферических тканях β-гидроксимасляная кислота окисляется до ацетоуксусной кислоты, а последняя активируется с образованием соответствующего КоА-эфира (ацетоацетил-КоА). Существует два ферментативных механизма активации ацетоуксусной кислоты. Первый путь -это использование АТФ и HS-КоА аналогично тому, как при активации жирных кислот: Реакция катализируется ферментом ацил-КоА-синтетазой, или тиокиназой. Второй путь активации - это перенос КоА от сукцинил-КоА на ацетоуксусную кислоту: Данная реакция катализируется ферментом сукцинил-КоА-ацетоацетат-трансферазой. Возможно, что биологически более важным является именно этот путь активации ацетоацетата. Образовавшийся в ходе этих реакций ацетоацетил-КоА подвергается далее тиолитическому расщеплению с образованием двух молекул ацетил-КоА: А уже ацетил-КоА, как это известно, окисляется в цикле трикарбоновых кислот (цикл Кребса) до СО2 и Н20. |