Физиология центральной нервной системы
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

 ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ Государственное образовательное учреждение высшего профессионального образования УЛЬЯНОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ИНСТИТУТ МЕДИЦИНЫ, ЭКОЛОГИИ И ФИЗИЧЕСКОЙ КУЛЬТУРЫ Н.Л. Михайлова, Л.С. Чемпалова ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ Учебное пособие Ульяновск 2010 УДК 612.82/.83(075.8) ББК 28.991.7я73 М 69 Рецензенты: д.б.н., профессор кафедры анатомии и физиологии человека Ульяновского государственного педагогического университета им. И.Н. Ульянова, заслуженный деятель науки РФ, академик РАЕН Л.Л. Каталымов; д.б.н., профессор, зав. кафедрой адаптивной физической культуры Ульяновского государственного университета М.В. Балыкин Михайлова, Н.Л. М 69 Физиология центральной нервной системы: учебное пособие / Н.Л. Михайлова, Л.С. Чемпалова. – Ульяновск: УлГУ, 2010. – 164 с. Учебное пособие написано на основе многолетнего опыта чтения лекций по физиологии центральной нервной системы для студентов специальностей «Лечебное дело», «Психология» и «Биология». Пособие состоит из двух частей: в первой части излагается материал по общей физиологии центральной нервной системы (ЦНС), во второй – по частной физиологии ЦНС. Изложение материала направлено на создание материальной основы для понимания механизмов интегративных процессов мозга и его роли в организации и регуляции функций. Учебное пособие предназначено для студентов высших учебных заведений, специализирующихся в различных областях медицины, физиологии, психологии, физкультуры и спорта, а также для всех интересующихся физиологией мозга. Может быть использовано для самостоятельной работы студентов. УДК 612.82/.83(075.8) ББК 28.991.7я73 © Михайлова Н.Л., Чемпалова Л.С., 2010 © Ульяновский государственный университет, 2010 ВВЕДЕНИЕ Центральная нервная система (ЦНС) в организме выполняет интегрирующую роль, объединяя в единое целое все ткани, органы и координируя их специфическую активность в составе целостных гомеостатических и поведенческих функциональных систем. Выполнение интегрирующей роли ЦНС осуществляет через свои функции. Основными функциями ЦНС являются: Управление деятельностью опорно-двигательного аппарата. ЦНС регулирует тонус мышц и посредством его распределения поддерживает естественную позу, а при нарушении восстанавливает ее, а также инициирует все виды двигательной активности. Регуляция работы внутренних органов. Осуществляется автономной нервной системой и эндокринными железами. Основная задача этой функции – поддержание гомеостаза (постоянства внутренней среды) в состоянии покоя и при различных видах деятельности. Обеспечение адаптивного поведения организмов в изменяющейся окружающей среде. Обеспечение высших психических функций: восприятие, внимание, эмоции, мышление, сознание, память. Язык как средство коммуникации, базирующееся на второй сигнальной системе. ЧАСТЬ 1 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ 1. Нейрон как структурно-функциональная единица нервной системы 1.1. Структура нервной клетки Нейроны, или нервные клетки, являются структурно-функциональ- ными единицами нервной системы. Несмотря на то, что эти клетки имеют те же самые гены, то же самое строение и тот же самый биохимический аппарат, что и другие клетки, они обладают и уникальными способностями, которые делают функцию мозга отличной от функции других органов. Важными особенностями нейронов является характерная форма, способность наружной мембраны генерировать нервные импульсы и наличие уникальной структуры, синапса, служащего для передачи информации от одного нейрона к другому. Нервные клетки чрезвычайно вариабельны по своему строению. В каждой из групп чувствительных, ассоциативных и двигательных нейронов имеется большое разнообразие форм, размеров тела клеток, величины и характера ветвления их отростков (рис. 1.1). Тело нейрона. По форме тела различают пирамидные, многоугольные, круглые и овальные клетки. На основании количества отходящих от тела клетки отростков все нейроны подразделяются на униполярные, биполярные и мультиполярные. Отростки могут отходить более или менее равномерно (радиально) от всей поверхности тела клетки либо концентрированно от одного из полюсов. Мультиполярные нейроны наиболее вариабельны по форме и имеют по нескольку отростков. Общепринято считать, что один из них – аксон (нейрит), который может начинаться как от тела клетки, так и от проксимальной части одного из дендритов. От тела биполярной клетки отходят два отростка. Тот из них, который направляется на периферию, принято считать дендритом, а центральный отросток – аксоном. Тела униполярных нейронов имеют овальную форму. От тела клетки отходит один крупный отросток, который на некотором расстоянии делится на два отростка: периферический и центральный. Форма клеточного тела целиком зависит от местоположения клетки в соответствующем участке нервной системы. На форму нервных клеток могут оказывать влияние соседствующие с ними кровеносные сосуды, пучки волокон или даже отдельные миелинизированные волокна крупного диаметра. Таким образом, одинаковые в функциональном отношении нервные клетки могут быть разными по форме.  Рис. 1.1. Нейрон зрительной коры кошки (микрофотография). На микрофотографии хорошо видны тело и дендриты нейрона (рисунок взят из книги: Мозг / под ред. П.В. Симонова. М.: Мир, 1984) Размеры нервных клеток колеблются в широких пределах. У разных по высоте организации животных и даже у одной и той же особи можно наблюдать примеры как очень мелких, так и очень крупных клеток. Так, диаметр клеток-зерен коры мозжечка равен приблизительно 5 мкм, а у моторных клеток головного и спинного мозга он достигает 70 мкм и более. Тела нервных клеток брюхоногих моллюсков хорошо различимы простым глазом (500-900 мкм). В нервной клетке существует прямая связь между массой сомы, величиной поверхности дендритов, калибром аксона, количеством коллатералей аксона и толщиной его миелинизированной оболочки. Было выяснено, что чем крупнее тела мотонейронов, тем длиннее у них аксон и больше дендритная поверхность нейрона. Форма нервных клеток определяется также комплексом их взаимосвязей с афферентными волокнами. Поэтому можно думать, что чем сложнее межнейронные связи каждого данного нейрона, тем сложнее его внешние очертания. Нейроны коры головного мозга характеризуются значительной вариабельностью форм. Подобно всем клеткам, нервные клетки отграничены сплошной плазматической мембраной от внешней среды. Тело нейрона содержит цитоплазму, ядро, есть микротрубочки и нейрофиламенты, органеллы и включения. В цитоплазме нейрона есть вещество Ниссля. Этот компонент цитоплазмы является определенным индикатором состояния нейрона, так как при изменении функционального состояния нейрона существенно изменяется вещество Ниссля. Основным компонентом вещества Ниссля является РНК. Количество РНК варьирует в зависимости от типа клеток и их размеров. Нервные клетки содержат также аппарат Гольджи, мультивезикулярные тела, лизосомы, пигменты (меланин или липофусцин). Меланин постоянно содержится в нейронах черной субстанции и голубого пятна. Присутствие меланина описано в дорсальном ядре блуждающего нерва, в ряде ядер ствола мозга и в симпатических нейронах. В тех клетках, где имеется меланин, мало липофусцина или он совершенно отсутствует. Липофусциновые гранулы, в отличие от меланина, начинают обнаруживаться в нейронах только с увеличением возраста. В нейронах обнаруживается большое количество митохондрий. Тело нейрона определяет процессы жизнедеятельности всей клетки и способность к регенерации ее отростков (рис. 1.2, 1.3). 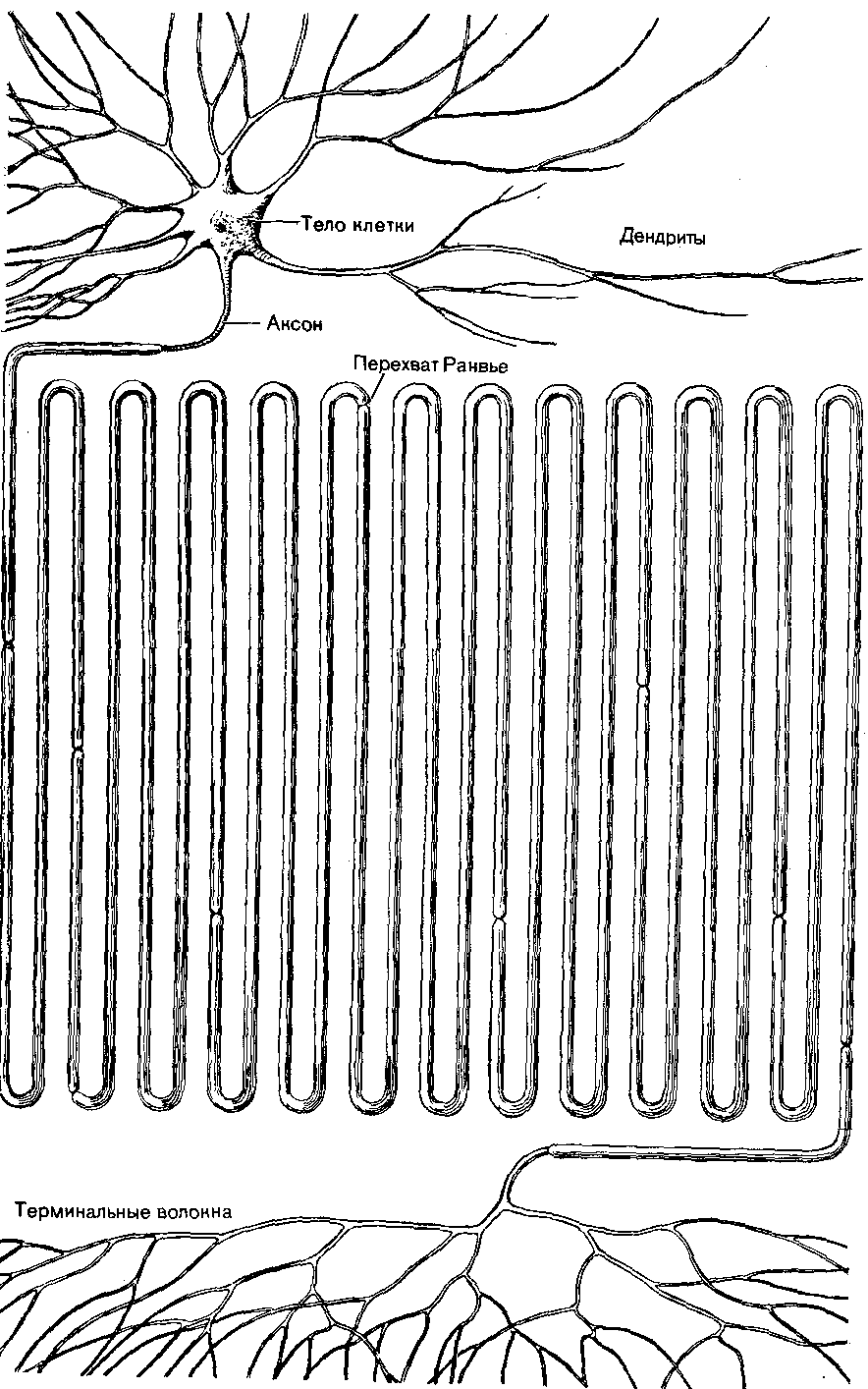 Рис. 1.2. Нейрон. Все части нейрона увеличены пропорционально (рисунок взят из книги: Мозг / под ред. П.В. Симонова. М.: Мир, 1984)  Рис. 1.3. Тело и дендриты нейрона (рисунок взят из книги: Мозг / под ред. П.В. Симонова. М.: Мир, 1984) Дендриты. Особенности, характерные для типичных дендритов и аксонов, приведены в таблице 1.1. Таблица 1.1 Особенности, характерные для типичных дендритов и аксонов
Терминали дендритов чувствительных нейронов образуют чувствительные окончания. Основной функцией дендритов является получение информации от других нейронов. Дендриты проводят информацию к телу клетки, а затем к аксонному холмику. Аксон. Аксоны образуют нервные волокна, по которым передается информация от нейрона к нейрону или к эффекторному органу. Совокупность аксонов образует нервы. Общепринято подразделение аксонов на три категории: А, В и С. Волокна группы А и В являются миелинизированными, а С – лишены миелиновой оболочки. Диаметр волокон группы А, которые составляют большинство коммуникаций центральной нервной системы, варьирует от 1 до 16 мкм, а скорость проведения импульсов равна их диаметру, умноженному на 6. Волокна типа А подразделяются на А, А, А, А. Волокна А, А, А имеют меньший диаметр, чем волокна А, меньшую скорость проведения и более длительный потенциал действия. Волокна А и А являются преимущественно чувствительными волокнами, которые проводят возбуждение от различных рецепторов в ЦНС. Волокна А – это волокна, которые проводят возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам. В-волокна являются характерными для преганглионарных аксонов вегетативной нервной системы. Скорость проведения 3-18 м/с, диаметр 1-3 мкм, продолжительность потенциала действия 1-2 мс, нет фазы следовой деполяризации, а есть длительная фаза гиперполяризации (более 100 мс). Диаметр С-волокон от 0,3 до 1,3 мкм, и скорость проведения импульсов в них несколько меньше величины диаметра, умноженного на 2, и равняется 0,5-3 м/с. Длительность потенциала действия этих волокон составляет 2 мс, отрицательный следовой потенциал равняется 50-80 мс, а положительный следовой потенциал – 300-1000 мс. Большинство С-волокон являются постганглионарными волокнами вегетативной нервной системы. В миелинизированных аксонах скорость проведения импульсов выше, чем в немиелизированных. Аксон содержит аксоплазму. У крупных нервных клеток ей принадлежит около 99% всей цитоплазмы нейрона. Цитоплазма аксонов содержит микротрубочки, нейрофиламенты, митохондрии, агранулярный эндоплазматический ретикулум, везикулы и мультивезикулярные тела. В разных частях аксона существенно меняются количественные отношения между этими элементами. У аксонов, как миелинизированных, так и немиелизированных, есть оболочка – аксолемма. В зоне синаптического контакта мембрана получает ряд дополнительных цитоплазматических соединений: плотные выступы, ленты, субсинаптическая сеть и др. Начальный участок аксона (от его начала до того места, где наступает сужение до диаметра аксона) носит название аксонного холмика. От этого места и появления миелиновой оболочки простирается начальный сегмент аксона. В немиелинизированных волокнах эта часть волокна определяется с трудом, а некоторые авторы считают, что начальный сегмент присущ только тем аксонам, которые покрыты миелиновой оболочкой. Он отсутствует, например, у клеток Пуркинье в мозжечке. В месте перехода аксонного холмика в начальный сегмент аксона под аксолеммой появляется характерный электронноплотный слой, состоящий из гранул и фибрилл, толщиной 15 нм. Этот слой не связан с плазматической мембраной, а отделен от нее промежутками до 8 нм. В начальном сегменте по сравнению с телом клетки резко уменьшается количество рибосом. Остальные компоненты цитоплазмы начального сегмента – нейрофиламенты, митохондрии, везикулы – переходят из аксонного холмика сюда, не изменяясь ни по внешнему виду, ни по взаиморасположению. На начальном сегменте аксона описаны аксо-аксональные синапсы. Часть аксона, покрытая миелиновой оболочкой, обладает только ей присущими функциональными свойствами, которые связаны с проведением нервных импульсов с большой скоростью и без декремента (затухания) на значительные расстояния. Миелин является продуктом жизнедеятельности нейроглии. Проксимальной границей у миелинизированного аксона служит начало миелиновой оболочки, а дистальной – утрата ее. Далее следуют более или менее длинные терминальные отделы аксона. В этой части аксона отсутствует гранулярный эндоплазматический ретикулум и очень редко встречаются рибосомы. Как в центральных отделах нервной системы, так и на периферии аксоны окружены отростками глиальных клеток. Миелинизированная оболочка имеет сложное строение. Ее толщина варьирует от долей до 10 мкм и более. Каждая из концентрически расположенных пластинок состоит из двух наружных плотных слоев, образующих главную плотную линию, и двух светлых бимолекулярных слоев липидов, разделенных промежуточной осмиофильной линией. Промежуточная линия аксонов периферической нервной системы представляет собой соединение наружных поверхностей плазматических мембран шванновской клетки. Каждый аксон сопровождается большим числом шванновских клеток. Место, где шванновские клетки граничат между собой, лишено миелина и называется перехватом Ранвье. Между длиной межперехватного участка и скоростью проведения нервных импульсов есть прямая зависимость. Перехваты Ранвье составляют сложную структуру миелинизированных волокон и играют важную функциональную роль в проведении нервного возбуждения. Протяженность перехвата Ранвье миелинизированных аксонов периферических нервов находится в пределах 0,4-0,8 мкм, в центральной нервной системе перехват Ранвье достигает 14 мкм. Длина перехватов довольно легко изменяется под действием различных веществ. В области перехватов, помимо отсутствия миелиновой оболочки, наблюдаются значительные изменения структуры нервного волокна. Диаметр крупных аксонов, например, уменьшается наполовину, мелкие аксоны изменяются меньше. Аксолемма имеет обычно неправильные контуры, и под ней лежит слой электронноплотного вещества. В перехвате Ранвье могут быть синаптические контакты как с прилежащими к аксону дендритами (аксо-дендритические), так и с другими аксонами. Коллатерали аксонов. С помощью коллатералей происходит распространение нервных импульсов на большее или меньшее число последующих нейронов. Аксоны могут делиться дихотомически, как, например, у зернистых клеток мозжечка. Очень часто встречается магистральный тип ветвления аксонов (пирамидные клетки коры мозга, корзинчатые клетки мозжечка). Коллатерали пирамидных нейронов могут быть возвратными, косоидущими и горизонтальными. Горизонтальные ответвления пирамид простираются иногда на 1-2 мм, объединяя пирамидные и звездчатые нейроны своего слоя. От горизонтально распространяющегося (в поперечном направлении к длинной оси извилины мозга) аксона корзинчатой клетки образуются многочисленные коллатерали, которые заканчиваются сплетениями на телах крупных пирамидных клеток. Подобные аппараты, так же как и окончания на клетках Реншоу в спинном мозге, являются субстратом для осуществления процессов торможения. Коллатерали аксонов могут служить источником образования замкнутых нейронных цепей. Так, в коре больших полушарий все пирамидные нейроны имеют коллатерали, которые принимают участие во внутрикорковых связях. За счет существования коллатералей обеспечивается в процессе ретроградной дегенерации сохранность нейрона в том случае, если повреждается основная ветвь его аксона. Терминали аксонов. К терминалям относятся дистальные участки аксонов. Они лишены миелиновой оболочки. Протяженность терминалей значительно варьирует. На светооптическом уровне показано, что терминали могут быть либо одиночными и иметь форму булавы, сетевидной пластинки, колечка, либо множественными и походить на кисть, чашевидную, моховидную структуру. Размер всех этих образований изменяется от 0,5 до 5 мкм и более. Тонкие разветвления аксонов в местах контакта с другими нервными элементами нередко имеют веретеновидные или бусинковидные расширения. Как показали электронно-микроскопические исследования, именно в этих участках имеются синаптические соединения. Одна и та же терминаль дает возможность одному аксону устанавливать контакт с множеством нейронов (например, параллельные волокна в коре головного мозга) (рис. 1.2). |