Физиология центральной нервной системы
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
12.2.2. Связи неокортекса В новой коре выделяют несколько типов эфферентных и афферентных связей. Эфферентными волокнами (кортико-фугальными) могут быть: 1) проекционные волокна к подкорковым образованиям (пути: кортико-спинальные, кортико-таламические, кортико-понтийные); 2) ассоциативные волокна, которые идут к одноименным и соседним областям коры одноименного полушария; 3) комиссуральные волокна, которые соединяют области коры обоих полушарий. Основными комиссурами являются мозолистое тело (corpus callosum) и передняя таламическая спайка. Мозолистое тело содержит очень много волокон. Например, у кошек на 1 мм2 приходится около 700 тысяч волокон. Афферентными волокнами (кортико-петальными) являются ассоциативные, комиссуральные и таламокортикальный путь – главный афферентный путь к коре от подкорковых образований. Афферентные волокна заканчиваются в основном в I-IV слоях коры. Исходя из этого, можно предположить, что в процессе обработки информации поверхностные слои ответственны главным образом за восприятие и обработку кортико-петальных сигналов. Особенное значение в этом процессе принадлежит IV слою коры. Тела важнейших эфферентных нейронов коры лежат преимущественно в более глубоких слоях V-VI. Их считают зоной начала эфферентных путей коры. 12.2.3. Специализация коры больших полушарий головного мозга Исходя из цитоархитектоники и характера связей коры, весь неокортекс был разделен на области и поля. Согласно классификации Бродмана, в коре головного мозга выделяют 11 областей и 52 поля (рис. 2.11). Так, в лобной доле (часть полушарий, расположенная кпереди от центральной борозды) выделяют прецентральную (моторная и премоторная) и собственно лобную (префронтальная) области. В пределах лобной области находятся поля 44 и 45 со специфическими для человека функциями. Так, в заднем отделе нижней лобной извилины находится область Брока, которая связана с моторной функцией речи. Собственно лобная или префронтальная кора имеет обширные связи (в большинстве ассоциативные) со всеми другими отделами коры мозга. Обширная разносторонне направленная система связей лобной области создает условия для интеграции информации из внешнего мира. 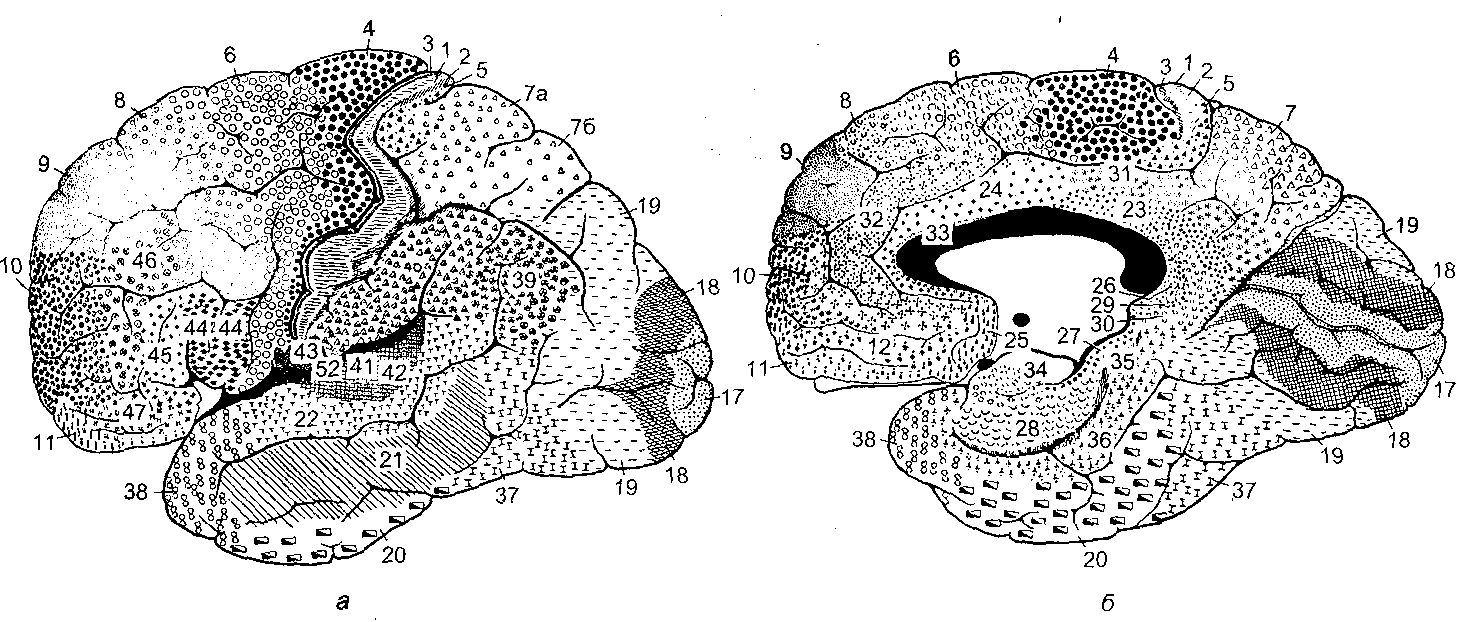 Рис. 2.11. Деление коры больших полушарий мозга на поля и области по Бродману. а – левое полушарие; б – правое полушарие Общими признаками так называемого лобного синдрома являются: отсутствие инициативы, апатия в сочетании с эйфорией, нарушение абстрактного мышления, неспособность усваивать новое, стереотипность в поведении и высказываниях, расторможенность влечений. При нарушении орбитальной лобной коры наступают глубокие нарушения личности. Прецентральная и постцентральная извилины входят в состав соматосенсорной коры. К этой области приурочена структурно-функциональная организация моторного и кожного анализатора. Основной функцией этой области является участие в регуляции двигательной активности. Поля 4 и 6 входят в состав прецентральной области. Поле 4 является центральной зоной моторной коры, а поле 6 – вторичной двигательной зоной (премоторная кора). Поле 4 является центральной зоной ядра двигательного анализатора. Моторная зона коры имеет четкую соматотопическую организацию. Постцентральная область содержит поля 1-3 и 43. 1-3 поля расположены на задней центральной извилине и парацентральной дольке. Поле 43 лежит горизонтально на роландовой покрышке. К постцентральной области приурочены анализаторы кожной и проприоцептивной чувствительности. Основной проекционной зоной кожной чувствительности являются поля 2 и 3. Известно, что в постцентральной области каждого полушария находятся соматотопические проекции противоположной стороны тела (рис. 2.12). Височная область имеет сложное строение и многообразные функции. На наружной поверхности височной доли располагается первичная проекционная слуховая зона – поля 41 и 42, а также поля 22 и частично 21, которые являются вторичной слуховой зоной, где осуществляется восприятие речи. Повреждение 21 и 22 поля ведет к нарушению фонематического слуха – синдрому сенсорной афазии. Большая часть 21 поля и поле 37 относятся к третичным ассоциативным полям, в которых происходит «перекрытие» зон различных сенсорных систем. Ассоциативные зоны височной коры особенно тесно связаны с образованиями лимбико-ретикуляр- ной системы. Двусторонняя деструкция височных долей у человека вызывает развитие амнестического синдрома, выраженные эмоциональные расстройства, нарушение влечений и высших психических функций. 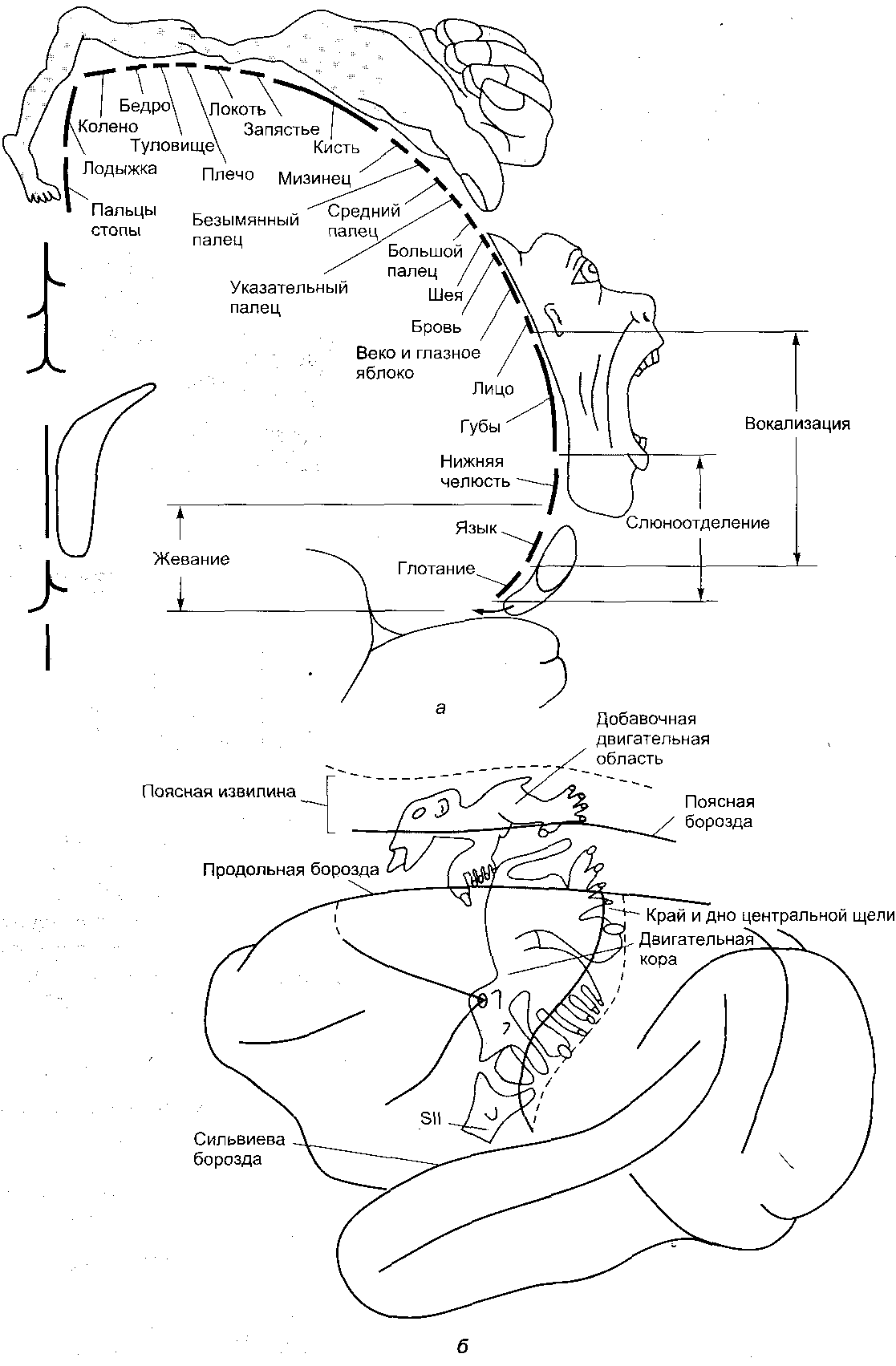 Рис. 2.12. Топографическая организация двигательной коры большого мозга (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004). а – латеральная поверхность; б – медиальная поверхность Теменная область расположена в задних отделах больших полушарий и относится к числу ассоциативных корковых зон. К верхним теменным полям относятся поля 5 и 7, а к нижним – поля 37, 39 и 40. Особенностью теменной коры является наличие в ней полисенсорных нейронов. Теменная кора, а точнее таламопариетальный комплекс, играет важную роль в межанализаторном синтезе сенсорной информации, необходимой для деятельности механизма высших гностических функций, а также для формирования системы «схема тела» (обобщенное представление человека о своем теле). Затылочная область является проекционным центральным звеном зрительной сенсорной системы. Она включает в себя поле 17 (первичное) и вторичные 18 и 19, кольцеобразно окружающие первичную зрительную зону. 12.2.4. Латеральная специализация больших полушарий головного мозга Мозг всех животных и человека билатерально симметричен – его правая и левая половины построены однотипно как по составу и количеству отдельных элементов, так и по общей архитектуре. У человека второй половины ХIХ века была обнаружена функциональная неравнозначность полушарий головного мозга. Впервые доминантность левого полушария по отношению к речевым функциям была обнаружена французским хирургом и антропологом Броком (1865). У людей, страдающих моторной афазией, им были обнаружены повреждения в левой лобной доле. Спустя десятилетие после открытий Брока Вернике показал, что поражение задней трети левой височной извилины левого полушария приводит к нарушению понимания речи и больной начинает воспринимать речь как нечленораздельные шумы. В этих случаях ведущими нарушениями являлись рецептивные функции речи. Поэтому Вернике обозначил весь синдром как синдром сенсорной афазии. В последующие годы в неврологических клиниках велось интенсивное изучение тех дефектов сложной деятельности мозга, которые возникали при очаговых поражениях одного из полушарий. На основе полученных фактов неврологи связали с деятельностью левого полушария не только речь, но и все высшие функции нервной системы – интеллект, сложные формы восприятия и деятельности. В результате сформировалась концепция тотального доминирования левого полушария человека. Левое полушарие головного мозга признавалось доминирующим не только в отношении языка, но и для концептуального мышления, определенных типов моторной деятельности и ориентации тела. Правое полушарие при этом считалось «субдоминантным», «малым», «немым», второстепенным, подчиненным левому, обслуживающим его. До 60-х годов ХХ века теория тотального доминирования левого полушария у человека почти безраздельно господствовала в неврологии. К началу 50-х годов были найдены функции, свойственные только правому полушарию. Стало очевидно, что правое полушарие вносит свой вклад в нервную деятельность. Сложилась новая концепция о функциональной специализации каждого из полушарий. К изучению функциональной асимметрии мозга подключились биологи, физиологи, биохимики и многие другие ученые. В результате на смену клиническим наблюдениям за больными стали разрабатываться новые методики исследований на здоровых людях и на животных. Бианки для исследований на животных предложил универсальный метод изучения – метод вызванных потенциалов. Этот метод позволил ему установить наличие функциональной асимметрии у разных видов животных. Он выявил моторную и сенсорную асимметрию, которая носила как видовой, так индивидуальный характер. Функциональная асимметрия перестала быть уникальной особенностью человеческого мозга. Бианки предположил, что деятельность правого и левого полушария строится на разных принципах: правое полушарие работает по принципу синтез – анализ, а левое – анализ – синтез. В основе функциональной асимметрии лежит принцип доминанты. На современном этапе изучения функциональной асимметрии мозга у здоровых людей применяются дихотические тесты, которые базируются на учете особенностей строения мозга. Известно, что правое ухо и правое поле зрения связаны более мощными путями с левым полушарием, а левое ухо и левое поле зрения – с правым полушарием. Если одновременно предъявлять разный материал правым и левым органам чувств, то полушария вступают в конкурентные отношения, и по особенностям восприятия удается судить о полушарной специализации. Кроме того, хорошо зарекомендовал себя метод временного выключения одного из полушарий электросудорожным ударом тока. В.Л. Деглин (2006) показал, что односторонний электрошок угнетает не весь мозг, а только то полушарие, над которым располагались электроды. Второе полушарие остается активным. Электроэнцефалограммы, записанные после шока, показывают поразительную картину – одно полушарие «спит» (регистрируются высокоамплитудные низкочастотные волны), а другое – бодрствует (регистрируется высокочастотная низкоамплитудная активность). В настоящее время накоплено достаточное количество фактов, которые дают возможность представить функциональные возможности каждого из полушарий. Так, у «левополушарного» человека (человек с выключенным правым полушарием) сохранена речь. Он охотнее и легче вступает в беседу, захватывает инициативу при разговоре, его словарь становится богаче и разнообразнее, ответы более развернутыми и детализированными. «Левополушарный» человек излишне многословен. Наряду с этим у него улучшается восприятие чужой речи. У «левополушарного» человека снижается порог обнаружения речи. Он быстрее и точнее повторяет слышимые слова. Вместе с тем, его речь теряет интонационную выразительность. Она становится монотонной, бесцветной, тусклой. Сам голос также изменяется. Он приобретает носовой, гнусавый оттенок либо становится неестественным, лающим. Такой дефект речи называется диспросодией, поскольку интонационно-голосовые компоненты речи называются просодическими («просодия» – мелодия). Нарушается также возможность воспринимать просодические компоненты речи собеседника. Выяснилось также, что «левополушарный» человек теряет способность понимать значение речевых интонаций. Он внимательно вслушивается, пытается расшифровать бессмысленные слоги, очень точно их повторяет, но сказать, с каким выражением (вопросительным, гневным и т.д.) они произнесены, не может. Не может он и отличить мужской голос от женского. Таким образом, наряду с сохранностью формального богатства речи, словарного и грамматического, наряду с увеличением речевой активности, обострением словесного слуха «левополушарный» человек потерял образность и конкретность речи. У него нарушается восприятие сложных звуков, музыкальных и зрительных образов. Оказалось, что «левополушарный» человек оказывается беспомощным при выполнении заданий, требующих ориентировки в наглядной образной ситуации, учета конкретных признаков предмета, восприятия ситуации в целом. У «левополушарного» человека улучшается настроение, он становится мягче, приветливее, веселее. Таким образом, у «левополушарного» человека сохранились или даже усилились те виды психической деятельности, которые лежат в основе абстрактного, теоретического мышления. Наблюдается положительный эмоциональный тонус. В отличие от «левополушарного», у «правополушарного» человека (у которого выключено левое полушарие) речевые возможности резко ограничены. Речь он понимает плохо. Порог обнаружения звуков речи у него повышен. В то же время голос «правополушарного» человека остается таким же. Не пострадал и слух на просодические компоненты речи, сохраняется ее интонационный рисунок. Такой человек даже лучше, чем обычно, различает мужские и женские голоса. Он хорошо узнает разнообразные несловесные звуки, мелодии песен, точно их воспроизводит. У «правополушарного» человека произошла перестройка восприятия – ухудшилось словесное восприятие и избирательно улучшились все виды образного мышления. У него произошел эмоциональный сдвиг в сторону отрицательных эмоций. Таким образом, имеющиеся факты показывают, что каждое из полушарий обладает своими уникальными возможностями при организации высших форм деятельности. Нормальная деятельность протекает при совместной деятельности полушарий. На самом деле ясно – два полушария, но мозг один. Оба полушария не независимы друг от друга. Между ними складываются сложные и противоречивые отношения. Эти отношения носят реципрокный характер (конкурентный) или комплементарный (дружеский). При комплементарном характере взаимодействия оба полушария дружески участвуют в работе мозга, дополняя способности каждого. При реципрокном взаимодействии каждое полушарие испытывает тормозные влияния со стороны партнера. Это является необходимым, чтобы адекватно реагировать на изменчивые обстоятельства и разнообразные ситуации. При этом приходится то сочетать способности правого и левого полушарий, то максимально использовать способности одного из них. Реципрокное взаимодействие позволяет всегда иметь наготове резервы, очень тонко и точно балансировать активность полушарий и тем самым соблюдать наиболее выгодное в данный момент соотношение образного и абстрактного мышления. Объединение способностей двух полушарий происходит при помощи комплементарного взаимодействия. Благодаря комплементарному взаимодействию соблюдается баланс между способностями каждого полушария. Таким образом, полноценная психика человека предполагает согласованную и уравновешенную работу обоих полушарий. К настоящему времени выяснено, что функциональная асимметрия мозга имеет под собой структурно-функциональную и биохимическую основу. Показано, что правое и левое полушария обладают разными связями со структурами ЦНС. Так, левое полушарие имеет более тесные связи с активирующими стволовыми структурами, а правое полушарие теснее связано с диэнцефалическим отделом, ответственным за вегетативную, гуморальную и эндокринную регуляцию. Кроме того, на данный момент установлено существование биохимической асимметрии. Например, вещества, которые характеризуются центральным активирующим эффектом, распределены в головном мозге асимметрично. Правое полушарие содержит более развитые адренергические системы, более чувствительно к алкоголю. У крыс норадреналина в миндалине, черной субстанции выше в правом или левом полушарии, что зависит от индивидуальных особенностей животных. Обнаружилось также, что в левом бледном шаре, базальных ядрах серотонина вырабатывается больше, чем в структурах правого полушария, а дофамина образуется больше в правой миндалине и левом полосатом теле, чем в противоположных одноименных структурах. Распределение рецепторов к различным биологически активным веществам также оказалось асимметричным. Так, распределение D2-рецепторов в полосатом теле асимметрично: у крыс-самцов их больше слева, а у крыс-самок – справа. Уровень гормонов в левых ядрах гипоталамуса выше, чем в правых. Таким образом, на данный момент установлено существование функциональной асимметрии всей центральной нервной системы, а не только больших полушарий. Появляются данные о том, что функционально асимметрично работают структуры правой и левой половин ЦНС не только при организации высших функций, но и при регуляции и организации вегетативных систем. Например, есть сведения о функционально асимметричном влиянии поясной извилины, лимбических ядер таламуса и гиппокампа на паттерн дыхания. Сегодня функциональная асимметрия мозга становится важнейшей проблемой науки о мозге. 13. Вегетативная (автономная) нервная система Согласно представлениям французского физиолога Биша (начало XIX века), функции животного организма были разделены на две группы: животной (анимальной, соматической) и вегетативной (растительной) жизни. Нервная система, обеспечивающая двигательные реакции скелетной мускулатуры, восприятие внешних раздражителей, получила название соматической нервной системы. Нервная система, которая обеспечивала рост, питание, размножение, обмен веществ, стала называться вегетативной нервной системой. Клод Бернар обозначил новый признак вегетативной нервной системы – иннервацию ею гладких мышц сосудов, внутренних органов, кожных образований. Из-за того, что гладкая мускулатура, иннервируемая вегетативной нервной системой, не подчиняется произвольному контролю, данная нервная система была названа Бернаром системой непроизвольной иннервации. Ленгли в дальнейшем эту непроизвольную часть нервной системы назвал автономной нервной системой. В настоящее время термин «автономная нервная система» употребляется чаще, чем «вегетативная нервная система». Ленгли разделил всю автономную нервную систему на два отдела: симпатический и парасимпатический. Соматическая и вегетативная нервная система различаются по конструкции рефлекторной дуги. Соматическая иннервация осуществляется однонейронным путем: тело эфферентного нейрона лежит в структурах ЦНС, а ее отросток (аксон), располагаясь на периферии, достигает исполнительного органа. В то же время непроизвольная вегетативная иннервация представлена двумя нейронами; один находится в ЦНС, а второй – в периферическом ганглии. 13.1. Рефлекторная дуга автономного рефлекса Рефлекторная дуга автономного рефлекса состоит из трех звеньев: чувствительного, вставочного и эффекторного. Чувствительное звено может быть образовано клетками спинномозговых или периферических ганглиев и может являться общим для автономной и соматической рефлекторных дуг. Периферические отростки чувствительных клеток разветвляются во внутренних органах, коже, стенках сосудов и т.д., центральные же синаптически контактируют со вставочными нейронами тех или других сегментов спинного мозга. Второе звено может быть в виде скопления нейронов в боковых рогах спинного мозга или в стволе мозга. Из боковых рогов спинного мозга отростки нейронов покидают спинной мозг в составе вентральных корешков, вступают в соматические нервные стволы и отсюда в виде белых соединительных ветвей направляются к узлам симпатического ствола. Здесь происходит синаптическое переключение части из них на эффекторные нейроны. Третье звено представлено нервными клетками, которые мигрировали из спинного мозга в один из периферических узлов. Узлы могут располагаться либо около позвоночника – это паравертебральные ганглии, либо в нервных сплетениях вблизи внутренних органах (превертебральные), либо в стенках внутренних органов (интрамуральные). Внутриорганные волокна и ганглии образуют сплетения, богатые нервными клетками, расположенные в мышечных стенках многих внутренних органов (сердце, бронхи, средняя и нижняя часть пищевода, желудок, кишечник, желчный пузырь, мочевой пузырь), а также в железах внешней и внутренней секреции. 13.2. Свойства вегетативных ганглиев Вегетативные ганглии играют важную роль в распределении и распространении нервных влияний на органы. Отмечено, что число нервных клеток в ганглиях в несколько раз превышает число преганглионарных волокон. Так, в верхнем шейном ганглии (симпатический) превышение составляет в 32 раза, а в ресничном узле (парасимпатический) – в 2 раза. Каждое из этих волокон сильно ветвится. Таким образом, расширяется зона влияния одного волокна. В ганглиях наблюдается явление конвергенции. Вместе с этим обнаруживается явление пространственной и временной суммации. У вегетативных ганглиев проявляются те же свойства, что и у соматических нервных центров. Поэтому ганглии вегетативной нервной системы иногда называют нервными центрами, вынесенными на периферию. Особенности возникновения возбуждения. В вегетативных ганглиях наблюдается большая длительность синаптической задержки (от 1,5 до 30 мс); большая длительность ВПСП, выраженная гиперполяризационная фаза ПД. Частота генерации нервного импульса невелика – 10-15 имп/с. Обнаруживается трансформация ритма. Если к ганглию подходит возбуждение с частотой импульсации свыше 100 имп/с, то наблюдается блокада проведения через синапс. Вероятно, свойства ганглиев и обеспечивают автономность вегетативных функций. Преганглионарные волокна принадлежат к типу В, их диаметр составляет 2-3,5 мкм, они тонкие и миелинизированные. Скорость распространения от 3 до 18 м/с. Потенциалы действия более длительные, чем у волокон соматических нейронов. Постганглионарные волокна относятся к типу С. Они являются немиелинизированными, тонкими, диаметром не более 2 мкм. Скорость распространения от 1 до 3 м/c. Для потенциала действия характерна длительная фаза гиперполяризации (до 3000 мс). 13.3. Симпатический и парасимпатический отделы автономной нервной системы В настоящее время всю автономную нервную систему подразделяют на симпатическую, парасимпатическую и метасимпатическую. Метасимпатическая – это нервная система, которая расположена внутри органа. Ее образует интрамуральный ганглий с системой нервных волокон. Высшим отделом симпатической и парасимпатической иннервации является гипоталамус. Медиаторами автономной нервной системы являются те же, что и в ЦНС: адреналин, норадреналин, ацетилхолин, вещество Р, гамма-амино- масляная кислота, гистамин, серотонин и др. Существуют определенные различия в морфофункциональной организации симпатического и парасимпатического отделов автономной иннервации. 1. Отличие проявляется в локализации центральных отделов симпатической и парасимпатической иннервации: центры симпатической нервной системы локализуются в основном в боковых рогах тораколюмбального отдела спинного мозга, а центры парасимпатической иннервации находятся в стволе мозга и в крестцовом отделе спинного мозга. Так, преганглионарные волокна парасимпатической нервной системы, снабжающие глазные мышцы и железы головы, покидают ствол мозга в составе трех пар черепно-мозговых нервов – III (глазодвигательный), VII (лицевой) и IX (языкоглоточный). К органам грудной и брюшной полости подходят в составе Х пары (блуждающий нерв) преганглионарные волокна, а к органам таза в составе тазовых нервов – парасимпатические волокна крестцового отдела спинного мозга. 2. Отличие проявляется также в типе ганглиев. Для симпатической нервной системы характерны паравертебральные и превертебральные ганглии, для парасимпатической – превертебральные и интрамуральные ганглии. 3. Различаются эти отделы и по длине пре- и постганглионарных волокон. Так, для симпатической нервной системы характерным является большей частью меньшая длина преганглионарных волокон, чем постганглионарных. Для парасимпатической нервной системы, наоборот, длина преганглионарных волокон может намного превышать длину постганглионарных. 4. Различие существует в медиаторах, которые осуществляют передачу в синапсах между иннервируемым органом и постганглионарным волокном. Медиатор в вегетативных ганглиях является единым для симпатического и парасимпатического отделов – это ацетилхолин, реализующий свое влияние через Н-холинорецепторы. В синапсах между постганглионарным волокном и эффекторным органом эти медиаторы разные. Для симпатической иннервации медиатором является адреналин и норадреналин, который связывается с α- и β-адренорецепторами на постсинаптической мембране эффекторного органа. Для парасимпатической иннервации это ацетилхолин, который реализует свое влияние через М-холинорецеп- торы на постсинаптической мебране эффекторного органа. 5. Физиологические эффекты при возбуждении симпатических или парасимпатических волокон у органов, имеющих двойную иннервацию, являются противоположными. Например, при возбуждении волокон блуждающего нерва, иннервирующих сердце, наблюдаются четыре отрицательных эффекта: инотропный (уменьшение силы сокращения), хронотропный (уменьшение частоты сокращения), батмотропный (уменьшение возбудимости) и дромотропный (уменьшение проводимости). При раздражении симпатических волокон, иннервирующих сердце, наблюдаются положительные эффекты. Блуждающий нерв для желудка является главным и секреторным, и моторным: активация его приводит к увеличению моторной и секреторной функции желудка. Активация симпатической нервной системы оказывает ингибирующий эффект на секреторную и моторную функцию желудка. Следует заметить, что эффекты, вызываемые раздражением симпатических волокон, зависят от рецепторов на постсинаптической мембране эффекторного органа. Так, при действии адреналина на α-рецепторы происходит сужение артерий и артериол скелетных мышц, а при действии адреналина на β-рецепторы в сердечной мышце сосуды в сердце расширяются. Механизм работы адренергических синапсов представлен на схемах, приведенных ниже (рис. 2.13-2.16). 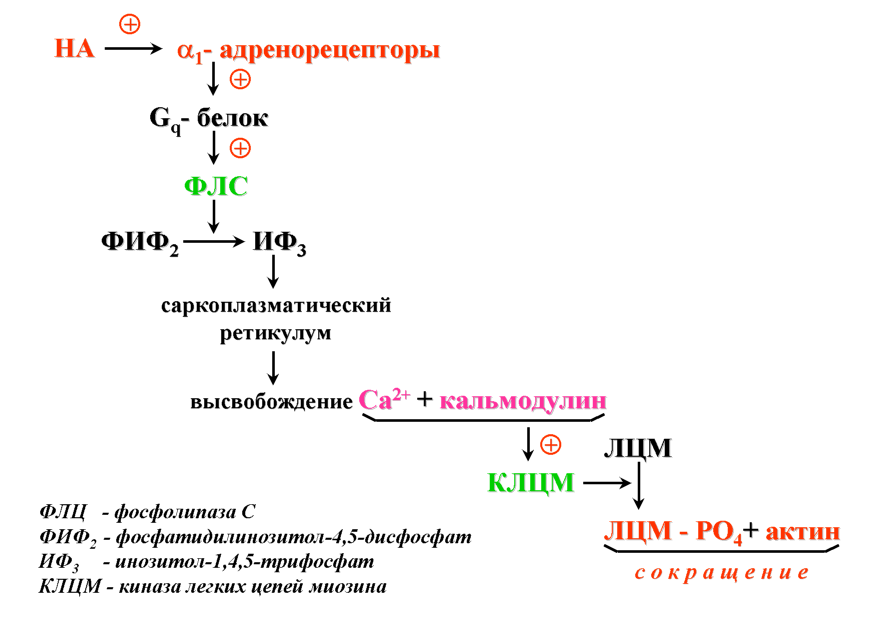  Основные эффекты: расширение зрачков (сокращение радиальной мышцы радужки); сужение кровеносных сосудов. Р 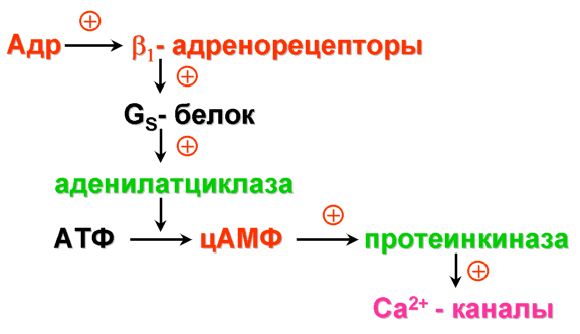  Основные эффекты: стимуляция деятельности сердца: повышение силы сокращений; повышение частоты сокращений; облегчение атриовентрикулярной проводимости; повышение автоматизма волокон проводящей системы. Р 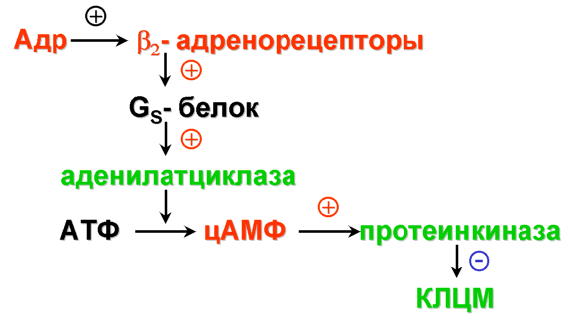  Основные эффекты: расширение кровеносных сосудов; расслабление гладких мышц бронхов; снижение тонуса и ритмической сократительной активности миометрия; гликогенолиз. Р   Основной эффект: сужение кровеносных сосудов Рис. 2.16. Механизм активации α2-адренорецепторов ЗАКЛЮЧЕНИЕ Центральная нервная система, состоящая из отдельных нейронов, связанных синаптическими контактами, представляет собой единое образование. Благодаря морфофункциональным особенностям нейронов, процессам торможения, различным медиаторным системам, по принципу доминанты в ЦНС создаются направленные потоки возбуждения, которые позволяют целостно и одновременно дифференцированно вовлекать мозг в любую из форм его активности. В работах И.П. Павлова и его учеников была высказана идея о динамической локализации функций. Эта идея получила дальнейшее развитие в работах физиологов, морфологов и клиницистов. В результате полученных фактов были разработаны принципы структурной и функциональной организации мозга и сформулирован эволюционный закон кортикализации функций. К настоящему времени убедительно доказано, что высшие функции мозга являются результатом системной деятельности мозга и в связи с этим поражение любого из отделов мозга может привести к распаду всей системы; таким образом, нарушение или выпадение функций не может непосредственно говорить о ее локализации. Динамический характер деятельности системы и межсистемной интеграции определяется особенностями поступающей афферентной импульсации, специфической реакцией организма и его внутренней активностью. Динамичность этих взаимоотношений имеет свои особенности на поведенческом, нейронном, синаптическом и молекулярном уровнях интеграции. С позиций системной организации функций в деятельности мозга выделяют различные функциональные системы и подсистемы (Анохин П.К., Лурия А.Р., Соколов Е.М., Адрианов О.С., Батуев А.С., Судаков К.В. и др.). Классический вариант интегративной деятельности мозга может быть представлен в виде взаимодействия трех основных функциональных блоков: 1) блока приема и переработки информации – сенсорные системы (анализаторы); 2) блока модуляции, активации нервной системы (лимбико-ретику- лярные системы мозга); 3) блока программирования, запуска и контроля поведенческих актов – моторные системы (двигательный анализатор). 1) Блок приема и контроля осуществляет процесс рецепции: в рецепторах сенсорных систем происходит трансформация энергии различных раздражителей в рецепторный и генераторный потенциалы; кодирования – на аксоне сенсорного нейрона происходит генерация потенциала действия и передача его по специфическим и неспецифическим путям в различные структуры ЦНС; перекодирования, анализа и синтеза. Этот процесс осуществляется в проекционных и ассоциативных областях коры, задействуются детекторные и гностические нейроны. 2) Модулирующие системы мозга регулируют тонус коры и подкорковых образований, оптимизируют уровень бодрствования в отношении выполняемой деятельности и обусловливают адекватный выбор поведения в соответствии с актуализируемой потребностью. Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные избирательные системы связей, программировать деятельность, осуществлять контроль над ней. Аппаратом, выполняющим функцию регулятора уровня бодрствования, а также осуществляющим избирательную модуляцию и актуализацию приоритета той или иной функции, является модулирующая система мозга. Ее часто называют лимбико-ретикулярный комплекс или восходящая активирующая система. К нервным образованиям этого аппарата относятся лимбическая и неспецифическая системы мозга с активирующими и инактивирующими структурами. Среди активирующих мозговых структур прежде всего выделяют ретикулярную формацию (РФ) среднего мозга, задний гипоталамус, синее пятно в нижних отделах ствола мозга. К инактивирующим структурам относят преоптическую область гипоталамуса, ядра шва в стволе мозга, фронтальную кору. Волокна ретикулярной формации, направляясь вверх, образуют модулирующие «входы» (аксо-дендритные синапсы) в вышерасположенных мозговых образованиях, включая старую и новую кору. От старой и новой коры берут начало нисходящие волокна, которые идут в обратном направлении к структурам гипоталамуса, среднего мозга и к более низким уровням мозгового ствола. Через нисходящие системы связей все нижележащие образования оказываются под управлением и контролем тех программ, которые возникают в коре головного мозга и для выполнения которых требуется модуляция активности и модификация состояния бодрствования. Таким образом, блок модуляции с его восходящими и нисходящими связями работает по принципу обратной связи как единый саморегулирующий аппарат, который обеспечивает изменения тонуса коры и вместе с тем сам находится под его контролем. Этот аппарат используется для пластичного приспособления организма к условиям среды. Он содержит два источника активации: внутренний и внешний. Первый связан с обменными процессами, обеспечивающими гомеостаз, а второй – с воздействиями внешней среды. Первый, например задний гипоталамус, обеспечивает поведенческую активацию. Второй источник активации связан с воздействиями раздражителей внешней среды. Часть непрерывного потока сенсорных сигналов, поставляемых в кору специфическими системами, по коллатералям поступает в РФ ствола. В аппарате восходящей РФ формируется механизм преобразования сенсорной информации в две формы активации: тоническую (генерализованную) и фазическую (локальную). Тоническая форма активации связана с функцией нижних стволовых отделов РФ. Она генерализованно, диффузно поддерживает определенный уровень возбудимости в коре и подкорковых образованиях. Фазическая форма активности связана с верхними отделами ствола мозга и прежде всего с неспецифической системой таламуса. Неспецифическая система таламуса локально и избирательно распределяет воздействия восходящей активации на подкорковые образования, старую и новую кору. Кроме того, установлено, что кора головного мозга наряду со специфическим функциональным вкладом оказывает «неспецифические» активирующие и тормозные влияния на нижележащие нервные образования. Корковые влияния, поступающие по нисходящим волокнам, представляют достаточно дифференцированную организацию и могут рассматриваться в качестве третьего источника активации. Наиболее обширные активирующие и инактивирующие избирательные влияния на ствол мозга исходят из лобных отделов коры. 3) Блок программирования, запуска и контроля поведенческих актов представляет собой сложноорганизованную, многоуровневую, построенную по принципу иерархии систему различных образований ЦНС. Замысел движения рождается в ассоциативных лобных отделах новой коры; при участии моторной коры, базальных ганглиев, таламуса и мозжечка создается программа движения. Создание программы движения осуществляется по принципу доминанты. При этом происходит отбор структур, которые будут необходимы для организации движения в конкретном поведенческом акте. Это делается путем формирования соответствующего тонуса двигательных ядер ствола и спинного мозга, одновременно устанавливается необходимое соответствие с тонусом двигательных ядер тонуса нервных центров, ответственных за вегетативное обеспечение поведения. Собственно движение осуществляется с участием двигательных единиц (ДЕ). Двигательная единица – мотонейрон спинного мозга и иннервируемая им группа мышечных волокон, образующих мышцу. Данный блок и все его узлы осуществляют свою деятельность на основе приема различного рода афферентаций (от зрительной сенсорной системы, вестибулярной, слуховой, тактильной, проприоцептивной, включая информацию от рецепторов растяжения мышц и т.д.). СПИСОК СОКРАЩЕНИЙ Адр – адреналин – медиатор и гормон мозгового слоя надпочечников. АТФ – аденилпирофосфорная кислота (аденозинтрифосфорная кислота) – нуклеотид, содержащий аденин, рибозу и 3 остатка фосфорной кислоты; универсальный переносчик и основной аккумулятор химической энергии в живых клетках, выделяющейся при переносе электронов в дыхательной цепи после окислительного расщепления органических веществ. цАМФ – циклическая форма аденозинмонофосфата, вторичный посредник в биохимических процессах клетки. АКТГ – кортикотропин, аденокортикотропный гормон – гормон передней доли гипофиза. Управляет ферментами синтеза глюкокортикоидной системы надпочечников. Обладает также липолитическим действием и усиливает пигментацию кожи. ВПСП – возбуждающий постсинаптический потенциал. ГАМК – гамма-аминомасляная кислота; тормозный медиатор. DОРА – дофамин – медиатор. ДЦ – дыхательный центр (продолговатого мозга). ИФ3 – инозитол-1, 4, 5-трифосфат. КЛЦМ – киназа легкой цепи миозина – фермент, принимающий участие в сокращении мышц. ЛО – локальный ответ. ЛЦМ – легкая цепь миозина. НА – норадреналин – медиатор. ПД – потенциал действия – быстрое колебание мембранного потенциала. РФ – ретикулярная формация. ТПСП – тормозный постсинаптический потенциал. ФИФ2 – фосфатидилинозитол-4, 5-дифосфат. ФЛС – фосфолипаза С. ЦНС – центральная нервная система. ЛИТЕРАТУРА Адрианов, О.С. О принципах организации интегративной деятельности мозга / О.С. Адрианов. – М.: Медицина, 1976. Батуев, А.С. Кортикальные механизмы интегративной деятельности мозга / А.С. Батуев. – Л.: Наука, 1978. Данилова, Н.Н. Психофизиология / Н.Н. Данилова. – М., 2000. Меркулова, Н.А. Дыхательный центр и регуляция его деятельности супрабульбарными структурами / Н.А. Меркулова, А.Н. Инюшкин, В.И. Беляков, Р.А. Зайнулин, Е.М. Инюшкина. – Самара, 2007. Механизмы деятельности мозга человека. Часть первая. Нейрофизиология человека / отв. ред. Н.П. Бехтерева. – Л.: Наука, 1988. Мозг / под ред. П.В. Симонова; пер. с англ. Н.Ю. Алексеенко. – М.: Мир, 1984. Общая физиология нервной системы / отв. ред. П.Г. Костюк. – Л.: Наука, 1979. Сафонов В.А. Человек в воздушном океане. М.: Национальное обозрение, 2006. Ухтомский, А.А. Доминанта / А.А. Ухтомский. – Л.: Наука, 1976. Физиология человека / под ред. Р. Шмидта и Г. Тевса. Т. 1. – М.: Мир, 1985. Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. – М.: Академия, 2004. Хомутов, А.Е. Физиология центральной нервной системы / А.Е. Хомутов. – Ростов н/Д: Феникс, 2006. Частная физиология нервной системы. Руководство по физиологии / отв. ред. П.Г. Костюк. – Л.: Наука, 1983. ТЕСТЫ ДЛЯ САМОКОНТРОЛЯ 1. В естественных условиях потенциал действия преимущественно возникает на мембране участка нейрона 1) сомы 2) пресинаптической 3) дендритах 4) начального сегмента аксона-аксонного холмика 2. Уровень деполяризации мембраны, при котором возникает потенциал действия, называется 1) субкритическим уровнем 2) нулевым уровнем 3) потенциалом покоя 4) критическим уровнем 3. Возбуждение в безмиелиновых нервных волокнах распространяется 1) скачкообразно, «перепрыгивая» через участки волокна, покрытые миелиновой оболочкой 2) в направлении движения аксоплазмы 3) непрерывно вдоль всей мембраны от возбужденного участка к расположенному рядом невозбужденному участку 4. Возбуждение в миелинизированных нервных волокнах распространяется 1) непрерывно вдоль всей мембраны от возбужденного участка к невозбужденному 2) электротонически и в обе стороны от места возникновения 3) в направлении движения аксоплазмы 4) скачкообразно, «перепрыгивая» через участки волокна, покрытые миелиновой оболочкой 5. Структурное образование, обеспечивающее передачу возбуждения с одной клетки на другую, носит название 1) нерв 2) аксонный холмик 3) перехват Ранвье 4) синапс 6. Мышечные волокна скелетных мышц иннервируются 1) нейронами симпатической системы 2) нейронами высших отделов головного мозга 3) мотонейронами УСТАНОВИТЕ СООТВЕТСТВИЕ 7. Виды потенциалов... представляют собой... А. Возбуждающий 1. Местную гиперполяризацию постсинаптический постсинаптической мембраны. потенциал 2. Распространяющуюся деполяризацию Б. Тормозный постсинаптической мембраны. постсинаптический 3. Местную деполяризацию потенциал постсинаптической мембраны. В. Потенциал 4. Местную деполяризацию постсинаптической концевой пластинки мембраны в нервно-мышечном синапсе. 8. Нервные волокна типа... проводят возбуждение со скоростью... А. А альфа 1. 3-18 м/с Б. В 2. 70-120 м/с В. С 3. 0,5-3 м/с ВЫБЕРИТЕ ОДИН ПРАВИЛЬНЫЙ ОТВЕТ 9. Роль синапсов ЦНС заключается в том, что они 1) являются местом возникновения возбуждения в ЦНС 2) формируют потенциал покоя нервной клетки 3) проводят токи покоя 4) передают возбуждение с нейрона на нейрон 10. В естественных условиях потенциал действия в нейроне возникает 1) в области дендритов 2) в синапсе 3) в соме нервной клетки 4) в начальном сегменте аксона 11. Интегральная деятельность нейрона заключается в 1) посттетанической потенциации 2) связи с другими нейронами посредством отростков 3) суммации всех постсинаптических потенциалов, возникающих на мембране нейрона 12. Явление изменения количества нервных импульсов в эфферентных волокнах рефлекторной дуги по сравнению с афферентными обусловлено 1) рефлекторным последействием 2) наличием доминантного очага возбуждения 3) посттетанической потенциацией 4) трансформацией ритма в нервном центре 13. С увеличением силы раздражителя время рефлекторной реакции 1) не меняется 2) увеличивается 3) уменьшается УСТАНОВИТЕ СООТВЕТСТВИЕ 14. Примером рефлекса... реакция А. Является 1. Сужение зрачка при яркой вспышке света. Б. Не является 2. Расширение зрачка при закапывании в глаз атропина (блокатора холинорецепторов). 15. Звено рефлекторной дуги... выполняет функции А. Рецепторное 1. Передает информацию о работе эффектора в кору головного мозга. Б. Афферентное 2. Центробежное проведение возбуждения от нервного центра к эффекторной структуре. В. Центральное 3. Центростремительное проведение возбуждения от рецепторов к нервному центру. Г. Эфферентное 4. Воспринимает энергию раздражителя и преобразует ее в нервный импульс. 5. Осуществляет анализ и синтез полученной информации. ВЫБЕРИТЕ ОДИН ПРАВИЛЬНЫЙ ОТВЕТ 16. При перерезке между красным ядром и ядром Дейтерса мышечный тонус 1) практически не изменяется 2) исчезает 3) значительно уменьшится 4) разгибателей становится выше 17. В спинном мозге замыкаются дуги всех перечисленных рефлексов, кроме 1) локтевого 2) подошвенного 3) мочеиспускательного 4) сгибательного 5) выпрямительного УСТАНОВИТЕ СООТВЕТСТВИЕ 18. В отделе ЦНС... располагаются... А. Продолговатом мозге 1. Центр речи. Б. Среднем мозге 2. Центры: сосудодвигательный, дыхательный, жевания, слюноотделения, глотания. В. Таламусе 3. Высшие подкорковые центры вегетативной нервной системы. Г. Гипоталамусе 4. Центры регуляции мышечного тонуса и непроизвольной координации движения. 5. Центры интеграции сенсорной информации от экстра- и интерорецепторов при передаче к коре большого мозга. ВЫБЕРИТЕ ОДИН ПРАВИЛЬНЫЙ ОТВЕТ 19. К специфическим тормозным нейронам относятся 1) нейроны черной субстанции и красного ядра среднего мозга 2) пирамидные клетки коры больших полушарий 3) нейроны ядра Дейтерса продолговатого мозга 4) клетки Пуркинье и Реншоу 20. По своему механизму постсинаптическое торможение может быть 1) только деполяризационным 2) только гиперполяризационным 3) и де-, и гиперполяризационным 21. По своему механизму пресинаптическое торможение может быть 1) и де-, и гиперполяризационным 2) только гиперполяризационным 3) только деполяризационным 22. После перерезки ниже продолговатого мозга мышечный тонус 1) практически не изменяется 2) исчезнет 3) усилится тонус разгибателей 4) значительно уменьшится УСТАНОВИТЕ СООТВЕТСТВИЕ 23. Эффекторным звеном рефлекса... могут быть... А. Вегетативного 1. Скелетные мышцы. Б. Соматического 2. Гладкие мышцы. 3. Секреторные железы пищеварительной системы. 4. Эпителиальные клетки кожи. 24. Эфферентные нейроны... располагаются... А. Симпатического отдела ЦНС 1. В интрамуральных ганглиях внутренних органов. Б. Парасимпатического отдела 2. В ядрах таламуса и гипоталамуса. ЦНС 3. В ганглиях симпатического ствола. 25. Тонические рефлексы... возникают при... А. Позы (положения) 1. Действии зрительных и слуховых сигналов. Б. Выпрямительные 2. Нарушении естественной позы. В. Статокинетические 3. Возбуждении вестибулярных рецепторов при изменении положения головы. 4. Возбуждении вестибулярных рецепторов при изменении скорости движения тела. 26. Мозжечковая недостаточность... проявляется... А. Астения 1. В нарушении походки. Б. Астазия 2. В треморе мышц. В. Атаксия 3. В ослаблении мышечного тонуса. 4. В слабости и быстрой утомляемости мышц. ВЫБЕРИТЕ ОДИН ПРАВИЛЬНЫЙ ОТВЕТ 27. Высшие центры регуляции вегетативных функций располагаются 1) в коре головного мозга 2) в таламусе 3) в продолговатом мозге 4) в гипоталамусе ОТВЕТЫ К ТЕСТАМ 1 – 4; 2 – 4; 3 – 3; 4 – 4; 5 – 4; 6 – 3; 7 – А3, Б1, В4; 8 – А2, Б1, В3; 9 – 4; 10 – 4; 11 – 3; 12 – 4; 13 – 3; 14 – А1, Б2; 15 – А4, Б1, 3, В5; 16 – 4; 17 – 5;18 – А2, Б4, В5, Г3; 19 – 4; 20 – 3; 21 – 3; 22 – 4; 23 – А3, 2; 24 – А3, Б1; 25 – А3, Б2, В4; 26 – А4, Б2, 3, В1; 27 – 4 ОГЛАВЛЕНИЕ Введение 3 Часть 1. Общая физиология центральной нервной системы 4 1. Нейрон как структурно-функциональная единица нервной системы 4 1.1. Структура нервной клетки 4 1.2. Функции нервных волокон 12 2. Физиология синапсов 19 2.1. Свойства химических синапсов 21 2.2. Медиаторы 22 2.3. Освобождение медиаторов 26 2.4. Механизм действия медиаторов 30 2.5. Механизм формирования возбуждающего постсинаптического потенциала 32 2.6. Механизм открытия ионного канала у метаботропных рецепторов 34 2.7. Тормозные постсинаптические потенциалы 35 2.8. Завершение синаптических процессов 38 2.9. Особенности возникновения возбуждения в нейроне 39 3. Торможение в центральной нервной системе 40 3.1. Постсинаптическое торможение 40 3.2. Пресинаптическое торможение 41 3.3. Тормозные нейронные сети 41 4. Нервный центр 42 4.1. Свойства нервных центров 43 5. Принципы координационной деятельности центральной нервной системы 44 5.1. Принцип рефлекса 44 5.2. Определение рефлекса 49 5.3. Классификация рефлексов 50 5.4. Принципы дивергенции, конвергенции, центральное облегчение, окклюзия 52 5.5. Принцип доминанты 53 6. Физиология клеток глии 58 6.1. Классификация глиальных клеток 59 6.2. Функции глиальных клеток 60 Часть 2. Частная физиология центральной нервной системы 63 7. Физиология спинного мозга 65 7.1. Морфофункциональная организация спинного мозга 65 7.2. Проводниковая функция спинного мозга 70 7.3. Рефлекторная деятельность спинного мозга 73 7.4. Спинальный шок 74 8. Физиология ствола мозга 75 8.1. Морфофункциональная организация ствола мозга 75 8.2. Ретикулярная формация ствола мозга 79 8.3. Рефлексы ствола мозга 97 9. Физиология мозжечка 101 9.1. Морфофункциональная организация мозжечка 101 9.2. Функции мозжечка 106 9.3. Мозжечок и вегетативные функции 108 10. Физиология промежуточного мозга 109 10.1. Таламус. Морфофункциональная организация. Функции 109 10.2. Гипоталамус. Морфофункциональная организация. Функции 116 11. Физиология лимбической системы 120 12. Физиология переднего мозга 121 12.1. Базальные ганглии. Морфофункциональная организация. Функции 121 12.2. Кора больших полушарий головного мозга 129 12.2.1. Функциональная гистология коры больших полушарий 129 12.2.2. Связи неокортекса 134 12.2.3. Специализация коры больших полушарий головного мозга 135 12.2.4. Латеральная специализация больших полушарий головного мозга 138 13. Вегетативная (автономная) нервная система 143 13.1. Рефлекторная дуга автономного рефлекса 143 13.2. Свойства вегетативных ганглиев 144 13.3. Симпатический и парасимпатический отделы автономной нервной системы 145 Заключение 152 Список сокращений 156 Литература 157 Тесты для самоконтроля 158 Учебное издание Михайлова Нина Леонидовна, кандидат биологических наук, доцент. Государственное образовательное учреждение Ульяновский государственный университет, медицинский факультет, кафедра физиологии и патофизиологии Чемпалова Любовь Сергеевна, кандидат биологических наук, доцент. Государственное образовательное учреждение Тольяттинский государственный университет, кафедра физического воспитания ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ Учебное пособие Оригинал-макет подготовлен Г.И. Петровой Подписано в печать 29.04.09. Формат 60х84/16. Усл. печ. л. 9,5. Тираж 500 экз. Заказ № 40/ Оригинал-макет подготовлен в Издательском центре Ульяновского государственного университета 432000, г. Ульяновск, ул. Л. Толстого, 42 Отпечатано с оригинал-макета в Издательском центре Ульяновского государственного университета 4 |