Физиология центральной нервной системы
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
12. Физиология переднего мозга В состав самого рострального отдела центральной нервной системы – переднего мозга входят базальные ганглии и кора больших полушарий головного мозга. 12.1. Базальные ганглии. Морфофункциональная организация. Функции Базальные ганглии являются структурами ядерного типа. Они расположены внутри больших полушарий между лобными долями и промежуточным мозгом. Базальные ганглии относятся к собственно подкорковым образованиям мозга в самом узком смысле этого понятия и включают в себя три парных образования: неостриатум, паллидум (бледный шар) и ограда (claustrum). Неостриатум состоит из двух ядер: хвостатого и скорлупы (n. caudatus, putamen). Неостриатум является филогенетически новой структурой. Наиболее отчетливо он представлен начиная с рептилий. Скорлупа и хвостатое ядро по происхождению, нейронному строению, ходу проводящих путей и нейрохимическому составу являются сходными. Оба ядра, по существу, представляют собой два тяжа серого вещества, разделенных почти на всем протяжении волокнами внутренней капсулы. Паллидум, бледный шар (globus pallidum), в отличие от неостриатума, является филогенетически более древним образованием; его гомолог обнаруживается уже у рыб. Ограда расположена между скорлупой и островковой корой. Филогенетически ограда является самым новым образованием. У ежей и некоторых грызунов ее еще нет. Морфофункциональные связи базальных ганглиев. Неостриатум образует связи с бледным шаром. Аксоны клеток неостриатума очень тонкие, до 1мкм, поэтому проведение возбуждения от неостриатума к паллидуму медленное. Стриапаллидарные волокна образуют в основном аксо-дендритные синапсы. Неостриатум оказывает двойственное влияние на нейроны паллидума – возбуждающее и тормозное. Неостриатум посылает прямые эфференты не только к паллидуму, но и к черной субстанции. Стрионигральные связи по своей природе моносинаптические и двусторонние. Большой интерес представляет обратная связь – от черной субстанции к неостриатуму. Считается, что аксоны нейронов черной субстанции, которые конвергируют к нейронам хвостатого ядра и к скорлупе, обеспечивают транспорт дофамина, синтезирующийся в нейронах черной субстанции. В неостриатуме он концентрируется в расширенных аксонных терминалях. Скорость транспорта дофамина по аксонам от черной субстанции к хвостатому ядру составляет приблизительно 0,8 мм в 1 час. Содержание дофамина в неостриатуме чрезвычайно велико. Имеются указания на то, что дофамина в неостриатуме млекопитающих в 6 раз больше, чем в паллидуме и передней части больших полушарий, в 19 раз больше, чем в мозжечке. Предполагается медиаторная роль этого амина в данной структуре. Кроме того, высказывается мнение о том, что дофамин активирует тормозные интернейроны неостриатума и таким образом подавляет деятельность его клеток. Выдвигается также предположение о том, что дофамин играет энергетическую роль в неостриатуме: через цАМФ он обеспечивает распад гликогена. Помимо теоретического интереса в изучении медиаторной и метаболической функции дофамина, особое значение приобретает участие дофамина в патологии. Было установлено, что у больных с двигательными расстройствами резко падает концентрация дофамина в обоих ядрах неостриатума – хвостатом и скорлупе. Стриаталамические связи. Неостриатум не имеет отчетливо выраженных моносинаптических связей с корой больших полушарий и с таламусом. Неостриатум осуществляет физиологическую связь с корой большого мозга и таламусом опосредованно, через бледный шар, который выступает в этом случае как неспецифическое ядро, как посредник в эфферентной импульсации хвостатого ядра и скорлупы. Постулируется замкнутый круг импульсации: неостриатум – паллидум – таламус – лобные доли – неостриатум. Этот круг носит название «каудатная петля». Ему придают большое значение в интеграции нервных процессов на высших уровнях мозга, в генезе синхронной активности коры, в регуляции сна и бодрствования. Кортикостриарные связи. Сейчас доказано, что почти от всех полей коры к хвостатому ядру и скорлупе конвергируют прямые волокна в составе внутренней капсулы и подмозолистого пучка. Наибольшее количество волокон идет к скорлупе и хвостатому ядру от передних отделов коры. Кортикостриарные волокна отличаются пространственной организацией. Топографически это проявляется в том, что передние области коры больших полушарий представлены в головке хвостатого ядра, а задние – в каудальном отделе хвостатого ядра (рис. 2.8). 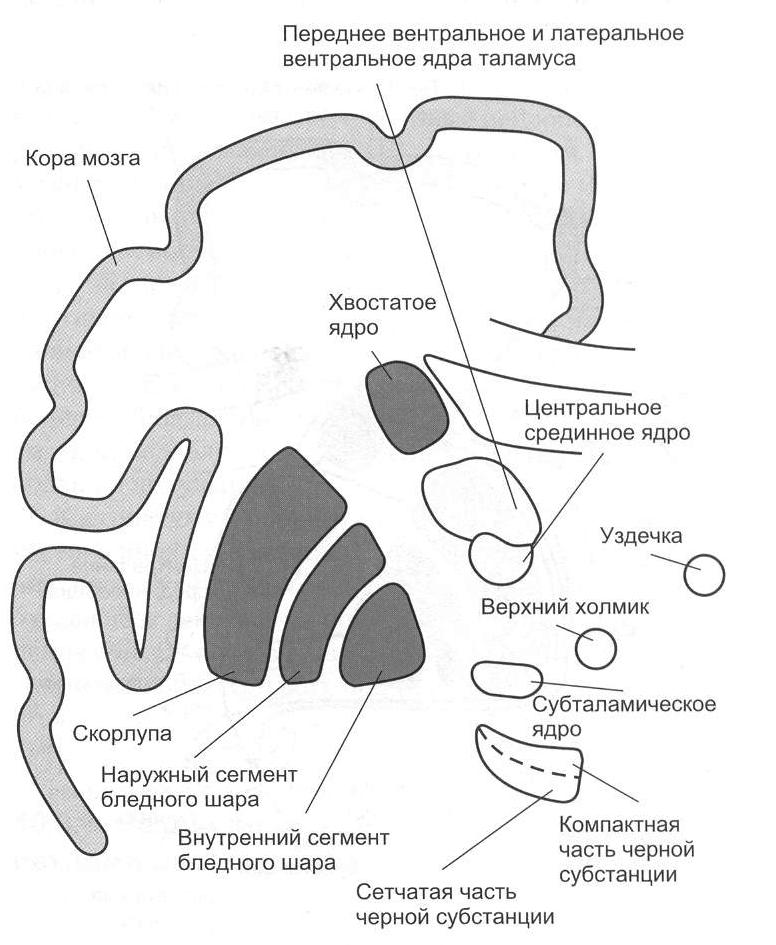 Рис. 2.8. Базальные ганглии и структуры, связанные с ними Функции базальных ганглиев. Этот комплекс ядер довольно широко включается в интегративную деятельность центральной нервной системы. Они играют определенную роль в ориентации животных в пространстве, запуске двигательного обеспечения пищевой мотивации, регуляции цикла бодрствование – сон. Неостриатум, паллидум, клауструм входят в программу осуществления условного рефлекса. Базальные ганглии и мозжечок являются равнозначными центрами, участвующими в программировании движений. Базальные ганглии могут иметь особое значение для осуществления стереотипных «червеобразных движений». Кроме того, каждая из структур обладает своими функциональными особенностями при вкладе в организацию движения. Неостриатум принимает участие в регуляции медленных движений, в которых преобладает тонический компонент. Паллидум дифференцирует характер движений: так, активность его нейронов у обезьян изменялась под влиянием толкательных движений, но эти же нейроны не реагировали на пронационные движения. Активность клауструма (у кошек) резко учащалась при болевых раздражениях. Отмечено также, что функциональные проявления базальных ганглиев определяются не столько связями отдельных ядер между собой, сколько связями каждого из них с другими структурами центральной нервной системы. Из этих структур наибольшее значение имеют неокортекс, неспецифические ядра таламуса, субталамическое ядро, черная субстанция, гипоталамус. На этом основании в настоящее время выделяют ряд функциональных петель базальных ганглиев. Скелетомоторная петля имеет входы от премоторной, моторной и соматосенсорной областей коры мозга. Основной поток информации идет через скорлупу, внутреннюю часть бледного шара или каудолатеральную область ретикулярной формации черной субстанции, затем через двигательные ядра таламуса и назад к шестому слою коры больших полушарий. При регистрации активности индивидуальных клеток скорлупы и бледного шара у обезьян, которые были обучены стандартным движениям, обнаружены четкие корреляции между этими движениями и активностью определенных нейронов. Наблюдается четкая топографическая организация: активность нейронов строго определенной области базальных ганглиев всегда соответствует специфическим движениям конкретных частей тела. Кроме того, во многих случаях наблюдается корреляция с особыми параметрами движения: силой, амплитудой или направлением движения. Регистрация активности клеток показала, что путь от стриатума через латеральную область ретикулярной формации черной субстанции управляет главным образом движением лица и рта. Окуломоторная (глазодвигательная) петля специализируется, вероятно, на регуляции движения глаз. Входные сигналы поступают от областей коры, контролирующих направление взгляда: фронтального глазного поля (поле 8) и каудальной части поля 7 теменной коры. Затем путь продолжается через хвостатое тело к дорсомедиальному сектору внутренней части бледного шара или к вентролатеральной области ретикулярной части черной субстанции. Затем идут связи к ядрам таламуса, которые дают проекции к фронтальному глазному полю. Аксоны нейронов сетчатой части черной субстанции раздваиваются, и одна ветвь идет к верхнему двухолмию среднего мозга, которое связано с движением глаз. Наблюдается положительная корреляция между активностью этих нейронов и саккадами (резкий перевод взгляда с одной точки на другую). Частота импульсации резко падает перед саккадой, что обусловлено тормозной стрианигральной связью (связью полосатого тела с черной субстанцией). Такое отключение тормозного выхода черной субстанции ведет к фазической активности таламуса или верхнего двухолмия. О полном пространственном разделении скелетомоторной и окуломоторной петель свидетельствует корреляция нейронной активности ретикулярной части черной субстанции с движениями либо глаз, либо рта, но никогда с теми и другими одновременно. К настоящему времени накоплены анатомические данные о существовании ряда «сложных петель», которые начинаются и заканчиваются в лобных ассоциативных областях коры (дорсолатеральной, префронтальной, латеральной орбитофронтальной, передней поясной), пройдя через ассоциативные ядра таламуса. В ходе филогенеза значительно возрастают размеры и значение корковых структур, стриатума и таламуса, участвующих в сложных петлях, так что у человека они становятся более обширными, чем двигательные. Однако функции сложных петель экспериментально еще не исследованы. Медиаторная система базальных ганглиев. Прохождение информации в описанных выше множественных параллельных трансстриальных функциональных петлях может облегчаться или подавляться модулирующими системами. Описано несколько модулирующих систем. Особого внимания среди них заслуживает дофаминергическая система. Дофаминергические нигростриальные пути (черная субстанция – полосатое тело) начинаются в сетчатой части черной субстанции. Содержащие дофамин нейроны обнаружены также поодиночке или группами вне черной субстанции, но поблизости от нее. Очень тонкие дофаминергические аксоны сильно ветвятся, образуя по всему стриатуму относительно диффузную сеть. Вдоль этих волокон находится множество небольших, заметных в световой микроскоп утолщений, называемых варикозами. На электронных микрофотографиях они идентифицируются как пресинаптические элементы. У нейронов сетчатой части черной субстанции довольно регулярная импульсация с частотой 1 Гц. Таким образом, каждую секунду импульс одной дофаминергической клетки вызывает высвобождение дофамина в многочисленных рассеянных по полосатому телу синапсах. Из-за своего диффузного строения дофаминергическая система не передает детализированную, топографически организованную информацию. Поэтому ее рассматривают как своего рода «ирригационную систему», модулирующую передачу информации по главному каналу. Так, было показано, что высвобождаемый в полосатом теле дофамин модулирует дофаминергическую кортикостриальную передачу (кора больших полушарий – полосатое тело). Восходящие дофаминергические волокна от среднего мозга направляются не только к стриатуму, но и к лимбическим структурам, к префронтальной коре. Аналогичное модулирующее влияние на базальные ганглии, возможно, оказывают серотонинергические волокна от ядер шва, норадренергические от голубого пятна, а также волокна с неизвестным медиатором от интраламинарных ядер таламуса и от миндалины; все они идут к полосатому телу. Кроме того, в базальных ганглиях содержится множество местных нейронов (интернейронов), модулирующих поток информации в трансстриатных петлях. К ним относятся холинергические нейроны полосатого тела и различные пептидергические нейроны. В течение длительного времени полосатое тело рассматривали как крупную однородную массу клеток, и лишь недавно была обнаружена его модульная организация. Окончания двух обширных систем афферентных волокон от коры больших полушарий и от ламинарных ядер таламуса образует здесь небольшие четко ограниченные центры. Анатомические эксперименты с дифференциальным окрашиванием волокон, относящихся к разным системам, показали, что в хвостатом ядре перемешаны скопления нервных окончаний от лобной и височной ассоциативной коры. Гистохимические методы дают аналогичную картину: разные медиаторы (глутамат, ГАМК, ацетилхолин, различные пептиды) обнаруживаются в пределах мелких, четко очерченных участков. Сейчас эти центры считаются независимыми компартментами, или микромодулями. Удалось проследить топографическую организацию в виде продольных колонок, идущих через весь стриатум. Таким же образом организованы проекции лобной и височной ассоциативной коры. С помощью микроэлектродного тестирования выявлены соматотопические продольные колонки, относящиеся к скелетомоторной петле. Например, в колонке верхней конечности, вероятно, собираются сигналы от премоторной, моторной и соматосенсорной областей коры. Нейроны в такой колонке объединены по сходству их соматотопических свойств. Тяжелые двигательные симптомы, сопровождающие нарушения в базальных ганглиях, можно разделить на две категории – гиперфункциональные и гипофункциональные. К гипофункциональным относят акинезию (отсутствие движений); к гиперфункциональным – ригидность (повышение мышечного тонуса), баллизм (крупномасштабный гиперкинез конечностей), атетоз (червеобразные движения), хорею (быстрые подергивания), тремор (дрожание). Симптомы второй категории являются результатом избыточной возбудимости двигательной системы, объясняемой растормаживанием. Гипофункция дофаминергической системы. Триада симптомов, характеризующая болезнь Паркинсона, наиболее распространенное заболевание базальных ганглиев, включает ригидность, тремор и акинезию. Вследствие мышечного гипертонуса усилены тонические (но не фазические) рефлексы растяжения; даже при медленном вытягивании конечности возникает восковая ригидность, часто периодического характера. Особенно сильно дрожат пальцы, а иногда губы и другие части тела. Акинезия проявляется рядом симптомов. Больному нелегко начать и завершить движение, временами произвольные движения могут «замирать». Становится чрезвычайно трудно выполнять одновременно два действия. Лицо больного невыразительно, как маска, модуляция речи ослаблена, во время ходьбы он не может взмахивать руками, идет мелкими шажками, обычно согнувшись вперед. У лиц, страдающих болезнью Паркинсона, дофаминергические нейроны черной субстанции отмирают. В начале ХХ века при вскрытиях таких больных было обнаружено, что у них отсутствует черный пигмент. Когда медиатор отмирающих нейронов был идентифицирован как дофамин, стало возможным объяснить потерю цвета гибелью самих нейронов и утратой дофамина. Утрата дофаминергических волокон и осуществляемого ими контроля нарушает нормальную деятельность двигательной системы. Регистрация активности нейронов полосатого тела показала, что их разряды возникают непосредственно перед началом движений определенного типа – медленных целенаправленных перемещений конечности из одной области пространства в другую. Когда человек, закрыв глаза, пытается рукой коснуться кончика носа, большая часть этого движения – перемещение руки из исходного положения почти до самого носа – осуществляется под контролем базальных ганглиев. Именно этот вид движений нарушается у больных паркинсонизмом. У животных с разрушенными в эксперименте дофаминергическими нейронами, аксоны которых оканчиваются в полосатом теле, наступает критический период, во время которого они как будто утрачивают способность начинать двигательные акты, даже такие важные, как еда и питье. Если животным предлагают пищу с сильным запахом, повышение сенсорной активации отчасти помогает им преодолеть это затруднение. Люди, страдающие болезнью Паркинсона, тоже могут временно преодолевать дефекты двигательной системы – в экстренных ситуациях, если, например, видят приближающийся автомобиль в тот момент, когда как раз собираются сойти с тротуара. У таких больных редко возникают трудности с речью или движением глаз. Это позволяет думать, что подобные функции осуществляются без участия дофамина. Больные паркинсонизмом должны принимать L-ДОФА в течение всей оставшейся жизни. Известно, что еще до начала применения L-ДОФА некоторого улучшения у больных удавалось добиться с помощью антагонистов ацетилхолина (производных атропина). В настоящее время считается, что в отсутствие дофаминергической системы происходит растормаживание холинергических нейронов. Согласно данной теории, в норме действие этих медиаторных систем тонко взаимосбалансировано. Гипофункция ГАМКергической и холинергической систем. Хорея, впервые описанная Гентингтоном, представляет собой наследственное дегенеративное заболевание базальных ганглиев, характеризующееся непроизвольными судорожными подергиваниями. При посмертном исследовании отмечается резкое уменьшение количества нейронов полосатого тела. Поражаются прежде всего синтезирующие ГАМК стриопаллидарные (полосатое тело – бледный шар) и стрионигральные (полосатое тело – черная субстанция) нейроны, а также местные холинергические клетки. Отсутствие осуществляемого стрионигральными нейронами торможения ведет к гиперактивности дофаминергических клеток. Это пример нарушения баланса, в данном случае между дофаминовой, ацетилхолиновой и ГАМК-системами. При гемибаллизме наблюдаются резкие непроизвольные размашистые движения с одной стороны тела, обычно как результат одностороннего повреждения субталамического ядра. Поскольку его ГАМКергические нейроны дают проекции к бледному шару, вероятно, такие движения обусловлены растормаживанием бледного шара. Однако попытки лечения этой болезни путем введения ГАМК или ее агонистов не принесли успеха. 12.2. Кора больших полушарий головного мозга 12.2.1. Функциональная гистология коры больших полушарий Кора головного мозга является высшим отделом центральной нервной системы. Она представляет собой тонкий слой нервной ткани, образующий множество складок. Общая площадь поверхности коры 2200 см2. Толщина коры колеблется от 1,3 до 4,5 мм. Объем коры равен примерно 600 см3. В состав коры головного мозга входит 109 – 1010 нейронов и еще большее число глиальных клеток (рис. 2.9). В пределах коры наблюдается чередование слоев, содержащих преимущественно тела нервных клеток, со слоями, образованными в основном их аксонами, и поэтому на свежем срезе кора головного мозга выглядит полосатой. На основании формы и расположения нервных клеток в коре с типичным строением можно выделить шесть слоев; некоторые из них подразделяются на два или более вторичных слоев. На основании строения коры выделяют следующие основные зоны: новую кору (неокортекс), старую кору (архикортекс), древнюю кору (палеокортекс) и межуточную кору (периархикортикальную и перипалеокортикальную). Самая обширная зона коры – неокортекс. Неокортекс занимает дорсальную и латеральную поверхность больших полушарий, в то время как палеокортекс находится на базальной и медиальной поверхности полушарий.  Рис. 2.9. Клеточный состав и слои коры головного мозга В неокортексе различают следующие слои: I. Молекулярный слой (плексиформный). В этом слое имеется множество волокон, образующих густое тангенциальное поверхностное сплетение, однако в нем мало клеток. Здесь содержатся в основном звездообразные мелкие клетки, которые осуществляют местную интеграцию деятельности эфферентных нейронов. II. Наружный зернистый слой. Содержит мелкие нейроны различной формы, которые имеют синаптические связи с нейронами молекулярного слоя на всем поперечнике коры. В его глубине располагаются малые пирамидные клетки. III. Наружный пирамидный слой. Этот слой состоит из пирамидных клеток малой и средней величины. Некоторые отделы коры в этом слое содержат крупные пирамидные клетки. Особенно много крупных пирамидных клеток в коре передней центральной извилины. Часть отростков этих клеток достигает первого слоя, участвуя в формировании тангенциального подслоя, другие погружаются в белое вещество полушарий мозга, поэтому III слой иногда обозначают как третичный ассоциативный. IV. Внутренний зернистый слой. Характеризуется рыхлым расположением мелких нейронов различной величины и формы с преобладанием звездчатых, имеющих дугообразные возвратные аксоны. Аксоны клеток проникают в выше- и нижележащие слои. Звездчатые клетки представляют систему переключений с афферентных на эфферентные нейроны III и IV слоев. В IV слое также образуется тангенциальная прослойка из нервных волокон. Поэтому иногда этот слой обозначается как вторичный проекционно-ассоциативный. Внутренний зернистый слой является местом окончания основной массы проекционных афферентных волокон. V. Внутренний пирамидный слой, или слой узловых клеток. Состоит в основном из средних и больших пирамидных клеток. Эти нейроны обладают длинными апикальными дендритами, простирающимися вплоть до молекулярного слоя, а также базальными дендритами, распространяющимися более или менее тангенциально по отношению к поверхности. Эти слои четко выражены в передней центральной извилине и незначительно – в других участках коры. Из этого слоя в основном формируются двигательные произвольные пути (проекционные эфферентные волокна). VI. Слой веретеновидных клеток (полиморфный, или мультиформный). В этом слое расположены преимущественно веретеновидные нейроны, которые имеют короткие извитые верхушечные дендриты, заканчивающиеся в V и IV слоях коры. Аксоны многих клеток слоя объединяются в возвратные волокна, проникая в V слой. Глубинная часть этого слоя переходит в белое вещество (рис. 2.10).  Рис. 2.10. Слоистое строение коры головного мозга Нужно заметить, что нейроны каждого коркового поля имеют свои особенности строения. Цитоархитектонические слои состоят из нервных и глиальных клеток (олигодендро-, астромакроглии) и многочисленных нервных волокон. Нервные волокна образуют плотные сплетения, называемые нейропилем. Нервные клетки по своей форме весьма разнообразны. Отмечают до 56 разновидностей клеток коры. Более обобщенно выделяют наиболее многочисленные пирамидные нейроны (гигантские Беца, крупные моторные, средние, мелкие), звездчатые и веретенообразные. Доля пирамидных клеток среди всех нейронов коры колеблется от 51 до 86%, звездчатых клеток – от 8 до 47%, веретенообразных нейронов – от 2 до 6% (рис. 2.9). Функционально в коре выделяют преимущественно возбуждающие нейроны: пирамидные, звездчатые, клетки Мартинотти (перевернутые пирамиды), глиеподобные и преимущественно тормозные: большие корзинчатые, малые корзинчатые, вертикально ориентированные, веретенообразные. Соединения между нейронами обеспечивают многочисленные синапсы и электротонические контакты. Большое значение в деятельности коры имеют шипиковые синапсы. Так, при развитии животных в условиях обогащенной раздражителями среды, по сравнению с сенсорной депривацией, происходит увеличение количества шипиков на дендритах. Умственная отсталость, снижение способности к обучению при хромосомных аберрациях у человека сопровождается уменьшением количества шипиков. Электротонические контакты осуществляются в коре в 20% случаев. Кроме того, в коре описаны несинаптические контакты между нейронами, функциональное назначение такого рода контактов остается еще неясным. В слоях I, II преимущественно имеются дендро-шипиковые контакты, в слоях III, IV – дендро-дендритические и сомато-дендритические, в слое V – сомато-сома- тические и дендро-дендритические. Американским физиологом В. Маунткаслом был выдвинут модульный принцип организации нейронов коры. Этот принцип базируется на трех отправных точках. 1. Кора головного мозга состоит из сложных многочисленных ансамблей, основная единица которых образована примерно сотней вертикально связанных нейронов всех слоев коры. Данный ансамбль называется мини-колонкой. В эти мини-колонки входят: а) нейроны, которые получают входные нейроны в основном от подкорковых структур, – например, от специфических сенсорных и моторных ядер таламуса; б) нейроны, получающие входные сигналы от других областей коры; в) все нейроны локальных сетей, образующих вертикальные клеточные колонки; г) клетки, передающие выходные сигналы от колонки назад к таламусу, другим областям коры, а иногда и клеткам лимбической системы. 2. Несколько таких сходных в своей основе простых вертикальных ансамблей могут объединяться с помощью межколоночных связей в более крупную единицу, перерабатывающую информацию, – модуль, или модульную колонку. Несмотря на различную плотность нейронов в слоях разных частей коры, общая структура и функции таких модульных колонок однотипны. Эти колонки различаются лишь по источнику получаемых ими входных сигналов и по мишеням, которым адресуются их выходные сигналы. 3. Маунткасл считает, что модули не только получают и перерабатывают информацию, но и совместно функционируют в составе обширных петель, по которым информация, выходя из колонок, передается другим кортикальным и субкортикальным мишеням, а затем возвращается обратно в кору. Эти петли обеспечивают упорядоченное поступление информации в кортикальные ансамбли. |