Физиология центральной нервной системы
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
Выпрямительные рефлексы обеспечивают возврат положения тела в нормальное – голова теменем вверх. Выделяют лабиринтные выпрямительные и шейные выпрямительные рефлексы. Для этих рефлексов необходимы красное ядро среднего мозга и вестибулярные ядра продолговатого мозга. Данные рефлексы возможны только у мезэнцефалического животного. Кроме того, мезэнцефалическое животное (с сохраненным средним мозгом) способно осуществлять ряд рефлексов, включающих определенные движения и возникающих также в результате движений, – статокинетические рефлексы. Многие из таких рефлексов обусловлены сигналами от вестибулярного аппарата: рефлексы повороты головы и глаз. Например, животное вращаем по часовой стрелке, голова животного вращается против часовой стрелки. К этой группе рефлексов относятся «лифтные рефлексы»: происходит увеличение тонуса мышц-разгибателей при линейном ускорении кверху и повышение тонуса сгибателей при ускорении книзу. К статокинетическим рефлексам относятся рефлекторные изменения тонуса мышц при сохранении равновесия тела при прыжках и беге, а также сложные рефлексы, благодаря которым кошка всегда падает на лапы. 9. Физиология мозжечка Мозжечок располагается у человека позади больших полушарий над продолговатым мозгом и мостом мозга. В эволюционном плане мозжечок представляет собой очень древнюю структуру. Впервые мозжечок появляется у круглоротых, что связано преимущественно с усложнением моторной координации. 9.1. Морфофункциональная организация мозжечка Анатомически в мозжечке можно выделить среднюю часть – червь, полушария мозжечка и боковые флокулонодулярные доли. Наиболее филогенетически молодой частью мозжечка является передняя часть задних долей полушарий мозжечка (неоцеребеллум). В полушариях мозжечка выделяют верхнюю поверхность – кору мозжечка и скопления нервных клеток – ядра мозжечка (рис. 2.5). 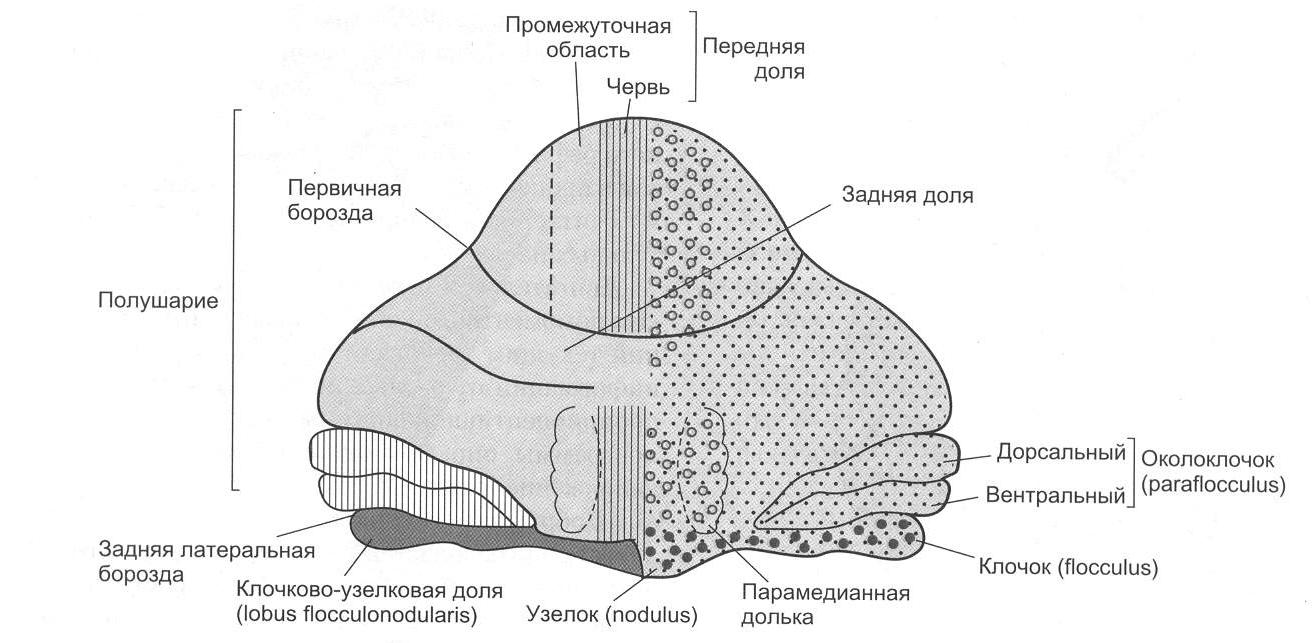 Рис. 2.5. Подразделение мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004) Нейронная организация коры мозжечка отличается исключительной упорядоченностью. В ней выделяют три слоя: I – поверхностный, или молекулярный слой. В этом слое располагаются дендритные разветвления грушевидных нейронов (клеток Пуркинье), параллельные волокна, которые являются аксонами вставочных нейронов. В нижнем слое молекулярного слоя находятся тела корзинчатых клеток, аксоны которых образуют синапсы с телами грушевидных нейронов, и некоторое число звездчатых клеток. II – ганглиозный слой. Здесь располагаются тела грушевидных нейронов или клеток Пуркинье. Клетки Пуркинье являются одними из наиболее сложно устроенных нейронов центральной нервной системы. Для этих нейронов примечательным является чрезвычайное ветвление дендритов. Дендритные отростки (дистальные части в особенности) обильно покрыты шипиками, что вместе с разветвлениями дендритов увеличивает площадь поверхностной мембраны, а следовательно, создает условия для размещения огромного числа синапсов. Один грушевидный нейрон может иметь до 200 000 синапсов. III – гранулярный слой. В этом слое находятся тела вставочных нейронов (клеток-зерен, или гранулярных клеток). Аксоны клеток-зерен поднимаются в молекулярный слой, где они Т-образно разветвляются. Здесь же находятся клетки Гольджи, аксоны которых также направляются в молекулярный слой (рис. 2.6). Важным для понимания функций мозжечка является изучение синаптических переключений в коре мозжечка и роли ее структур в деятельности ядер мозжечка. Изучением межклеточных связей в коре мозжечка занимался Экклс (рис. 2.7). 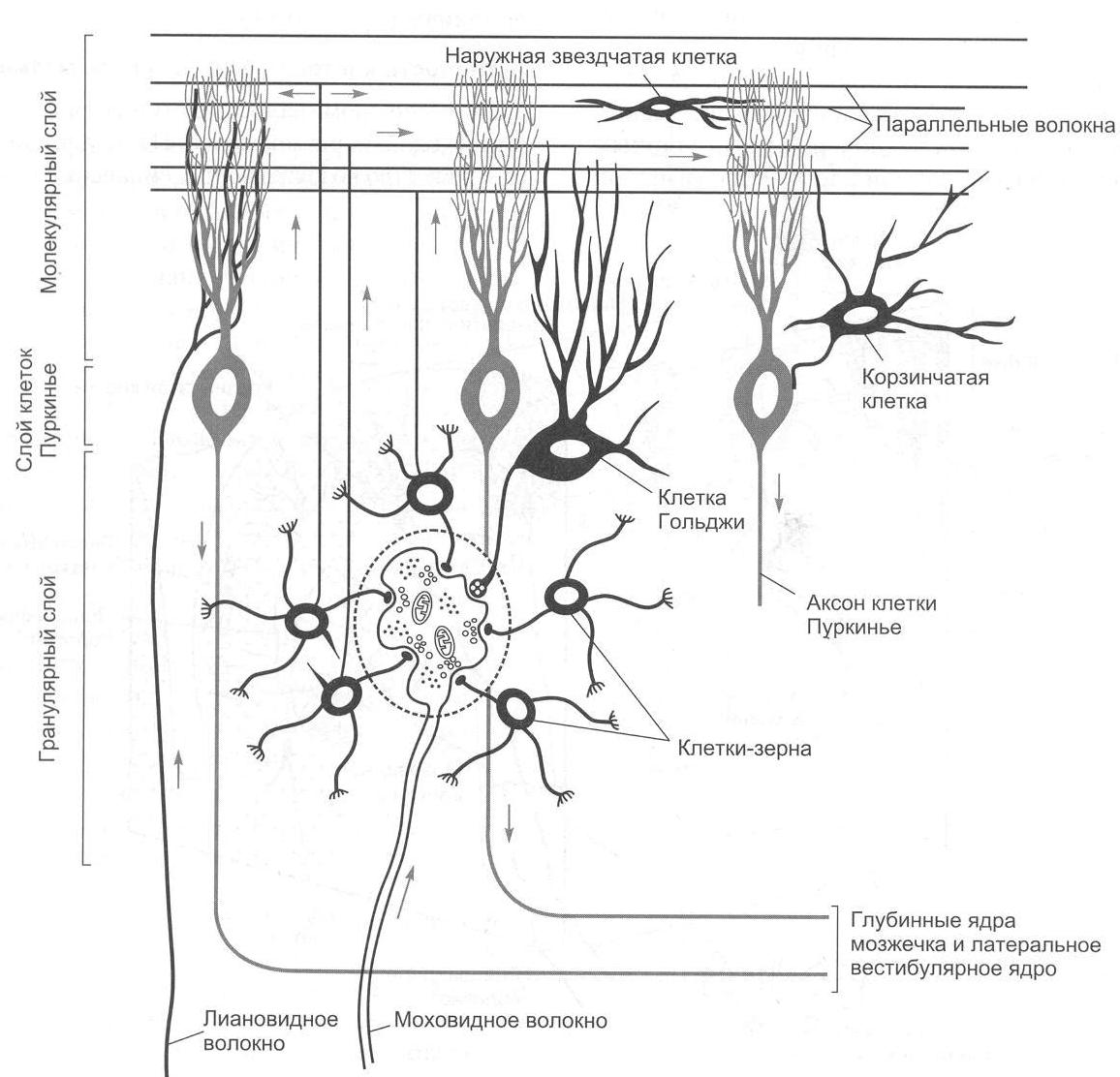 Рис. 2.6. Схема среза коры мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004) 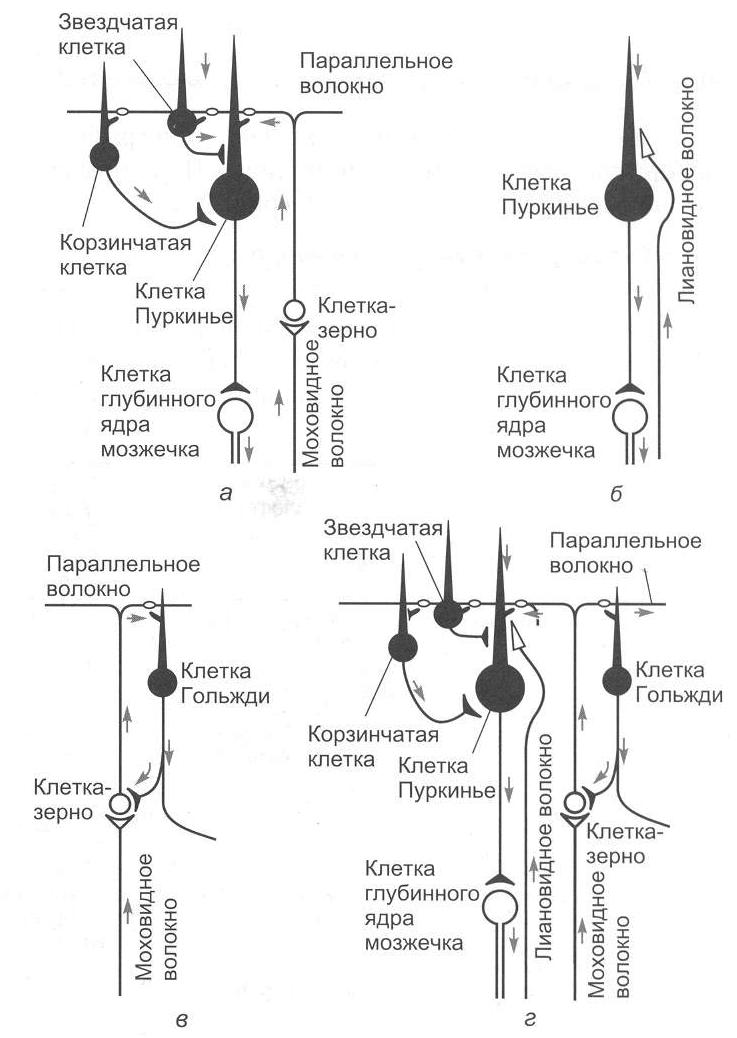 Рис. 2.7. Нейронные цепи коры мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004) Вход в мозжечок происходит через кору мозжечка с помощью двух типов волокон – лиановидных (лазающих) и мшистых (моховидных). Лиановидные волокна образуют многочисленные возбуждающие синапсы на дендритах клеток Пуркинье. При этом одного импульса достаточно для того, чтобы клетка Пуркинье ответила целым разрядом. Мшистые волокна возбуждают клетки-зерна, а те через параллельные волокна оказывают возбуждающее действие на все остальные нейроны, которые, однако, всегда являются тормозными: клетки Гольджи тормозят клетки-зерна по принципу обратной связи, а разряды клеток Пуркинье, воникающие под действием мшистых или лиановидных волокон, приводят к торможению ядер мозжечка. Таким образом, все клетки, нейроны, тела которых лежат в коре мозжечка, кроме клеток-зерен, выполняют тормозные функции. Ни в одном отделе центральной нервной системы нет такого преобладания торможения над возбуждением. Нейроны Пуркинье обладают активностью покоя, который обусловливает тоническое торможение ядер мозжечка. При увеличении активности клеток Пуркинье вследствие возбуждения мшистых или лиановидных волокон торможение ядер мозжечка усиливается; если же происходит торможение клеток Пуркинье (прямое – звездчатыми, непрямое – клетками Гольджи), то ядра мозжечка растормаживаются. Поскольку любое возбуждение, которое поступило в мозжечок, пройдя самое большое, через два синапса, превращается в торможение, уже через 100 мс это возбуждение угасает, и область мозжечка, в которое оно пришло, вновь становится готовой принять новую информацию. Возможно, такое автоматическое стирание информации играет важную роль в связи с участием мозжечка в организации быстрых движений. В состав мозжечка входят следующие ядра: ядро Шатра, вставочное ядро (у человека это ядро состоит из пробкового и шаровидного ядер) и зубчатое ядро. Афферентные и эфферентные связи мозжечка. Афферентные и эфферентные связи мозжечка очень многочисленные. Мозжечок получает афферентные связи от вестибулярных ядер, соматосенсорные пути, главным образом от спинного мозга, нисходящие пути – в основном от коры головного мозга. От спинного мозга к мозжечку подходят два спиномозжечковых пути – вентральный и дорсальный (Говерса и Кларка). В настоящее время выделяют приблизительно еще 10 трактов. Примерно половина из них, включая пути Говерса и Кларка, входят в мозжечок в виде мшистых волокон. Остальные же пути представляют собой спинооливарные тракты, которые, переключаясь на нейроны в оливах, посылают лиановидные волокна к коре мозжечка. Все области коры больших полушарий, а также другие отделы головного мозга посылают нисходящие пути к мозжечку. Большинство этих путей образуют синапсы в ядрах варолиева моста, волокна от которых идут к нео- или понтоцеребеллуму. Сигналы от двигательных зон коры больших полушарий поступают главным образом в промежуточную часть мозжечка, а импульсы от остальных корковых участков – к его полушариям. Эфферентные связи. Кора червя мозжечка посылает пути к ядру Шатра, средняя часть коры мозжечка (латеральнее червя) – к вставочному ядру, а кора полушарий мозжечка – к зубчатому ядру. Пути от каждого из этих ядер поступают к различным образованиям ствола мозга и больших полушарий. Таким образом, ядра мозжечка являются выходными структурами мозжечка. 9.2. Функции мозжечка Изучение мозжечка происходило в основном в связи с его участием в организации движения. В результате сложилось мнение, что основное значение мозжечка состоит в дополнении и коррекции деятельности остальных двигательных центров. Мозжечок отвечает за регуляцию позы и мышечного тонуса, за исправление (при необходимости) медленных целенаправленных движений в ходе их выполнения и координацию этих движений с рефлексами поддержания позы, правильное выполнение быстрых целенаправленных движений, команда к которым поступает от коры больших полушарий головного мозга. При осуществлении своих двигательных функций задействуются различные участки коры мозжечка, его ядра и ядра ствола мозга. Так, при взаимной координации позных и целенаправленных движений, при коррекции выполняющихся движений (особенно при выполнении незаученных или редко выполняющихся движений) принимает участие кора промежуточной части мозжечка, вставочное ядро мозжечка, красное ядро среднего мозга. Предполагается, что вставочное ядро через ядра таламуса связывается с двигательной корой и через рубро-спинальный и кортико-спинальный тракты влияет на мотонейроны спинного мозга. Связи коры полушарий мозжечка, зубчатого ядра мозжечка, красного ядра ствола мозга, таламуса, двигательной коры и ассоциативной коры мозга используются главным образом для генерации и осуществления быстрых баллистических целенаправленных движений. К этим отделам мозжечка, в отличие от остальных его частей, сигналы поступают не непосредственно от периферических органов, а от ассоциативных областей коры головного мозга. Замысел действия превращается в полушариях мозжечка и в его соответствующих ядрах в программу действия. Червь мозжечка через ядро Шатра и ядро Дейтерса принимает участие в регуляции позы. Нарушение функций мозжечка. Нарушение функций мозжечка проявляется в различных видах нарушения движений. 1. Асинергия – невозможность посылать должное количество нервных импульсов к различным мышцам, которые выполняют движение. При этом отдельные компоненты двигательной программы выполняются не одновременно, а скорее последовательно (распад движения). Асинергия приводит к следующим феноменам: 1.1. Дисметрия – движения выполняются в избыточном или недостаточном объеме, а затем наступает их чрезмерная компенсация. 1.2. Церебеллярная атаксия – у таких больных наблюдается неверная походка с широко расставленными ногами и избыточными движениями. Становится невозможным выполнение быстрой последовательности движений (адиадохокинез, или дисдиадохокинез). 2. Интенционный тремор – тремор, который отсутствует в покое, но возникает при движении. 3. Гипотония – снижение мышечного тонуса. Этот симптом часто сопровождается слабостью и быстрой утомляемостью мышц (связано главным образом с повреждениями полушарий мозжечка). 4. Нистагм глаз. 5. Головокружение. 6. Дефекты речи. Типичным проявлением поражений мозжечка служит триада Шарко: нистагм, интенционный тремор и скандированная речь. 9.3. Мозжечок и вегетативные функции Факты, указывающие на причастность мозжечка к регуляции деятельности внутренних органов, стали накапливаться сразу же, как только было начато экспериментальное исследование его функций. Клод Бернар (1858) наблюдал у голубей после укола в мозжечок расстройства моторной и секреторной функции пищеварительного тракта и нарушения углеводного обмена диабетического характера. Экхард (1872) описал явления сахарного диабета у собак и кроликов после воздействия электрического тока на червь мозжечка. Им были отмечены изменения со стороны сердца и дыхания. Овсянников (1875) подтвердил в опытах на кроликах факты, полученные Экхардом, но вегетативные расстройства он связывал с повреждением других областей мозга, а не мозжечка. Лючиани (1893) наблюдал расстройства в деятельности внутренних органов, а также трофические поражения у собак при удалении у них мозжечка. Так, в первый период у безмозжечковых собак отмечалось отсутствие аппетита, нарушение деятельности пищеварительного тракта в виде затруднения захватывания пищи и глотания, диареи. Изменялся обмен веществ. Показателями нарушений обмена веществ служили полиурия и гликозурия, исхудание, иногда ожирение. К исходу 10-20 дней расстройства деятельности пищеварительного тракта проходили или уменьшались, но обнаруживались дистрофические явления в виде облысения и потери блеска волос, эритематозных и экзематозных сыпей, маразма, уменьшалось заживление ран и т.д. Но описанные вегетативные расстройства Лючиани трактовал как неспецифические реакции организма на повреждение нервной системы вообще, а роль мозжечка он видел только в организации движения. Авторитет Лючиани был очень велик, что мешало, несмотря на многочисленные факты, оценить роль мозжечка в регуляции вегетативных функций. Принципиально новой в представлении о функциях мозжечка, в том числе и в отношении вегетативных процессов, по праву следует считать систему взглядов отечественного физиолога Л.А. Орбели. На основании обобщения многочисленных работ своей лаборатории Л.А. Орбели (1949) пришел к выводу, что роль мозжечка нельзя ограничивать какой-либо одной сферой деятельности организма. По мнению Орбели, назначение мозжечка проявляется, с одной стороны, «… в мобилизации ресурсов рефлекторных дуг в целом и их отдельных звеньев», а с другой стороны, «… в стабилизации их, удержании в определенных рамках возбудимости, лабильности, эффективности, генерализованности…» (Орбели Л.А., 1949). Согласно Орбели, мозжечок выполняет функцию стабилизатора нормальных взаимоотношений между различными отделами нервной системы. И хотя ни одна из функций организма не выпадает после удаления мозжечка, тем не менее они претерпевают существенные изменения количественного порядка в ту или иную сторону. Такая концепция послужила основой учения об адаптационно-трофической роли мозжечка в организме. Работы Асратяна (1941) показали, что при раздражении мозжечка электрическим током умеренной силы заметных изменений в вегетативных реакциях не происходило, но получались сдвиги в возбудимости соответствующих вегетативных центров. Именно в этом и проявляется приспособительное, адаптационно-трофическое влияние мозжечка на вегетативные процессы. Братусь (1969) показал с помощью метода вызванных потенциалов функциональные связи мозжечка с внутренними органами. Он получил вызванные потенциалы (ВП) коры мозжечка при раздражении чревных нервов, блуждающих нервов, нервных ветвей желудка и кишечника, механорецепторов желудка и кишечника. Эти работы подтвердили правильность концепции Л.А. Орбели. Современные ученые относят мозжечок к интегративным системам мозга. Это нисколько не противоречит взглядам Орбели. Мозжечок как интегративная система мозга принимает участие в составлении двигательных программ при организации функциональных систем целенаправленного поведения. При этом в его задачу входит отобрать соответствующие двигательные центры (по принципу доминанты) и привести в соответствие с двигательной программой состояние тех нервных центров, которые будут создавать вегетативное обеспечение, адекватное уровню активности организма. 10. Физиология промежуточного мозга 10.1. Таламус. Морфофункциональная организация. Функции Таламус, или зрительный бугор, является составной частью промежуточного мозга. Он занимает центральное место между большими полушариями. Особая локализация таламуса, его теснейшие связи с корой больших полушарий головного мозга и афферентными системами обусловливают особую функциональную роль этого образования. Как отмечал Уолкер (1964), «… в таламусе, этой огромной нейронной массе, лежит ключ к тайнам церебральной коры…». Таламус представляет собой массивное парное образование, имеющее яйцевидную форму, длинная ось которого ориентирована в дорсовентральном направлении. Медиальная поверхность таламуса образует стенку III желудочка, верхняя является дном бокового желудочка, наружная прилегает к внутренней капсуле, а нижняя переходит в гипоталамическую область. Таламус является ядерным образованием. В нем различают до 40 пар ядер. В настоящее время существует множество делений ядер таламуса на группы, в основе которых лежат различные принципы. Согласно Уолкеру (1966), а также Смирнову (1972), по топографическому признаку все ядра делят на 6 групп. 1. Передняя группа ядер включает ядра, составляющие передний бугор таламуса: переднее дорсальное (n. AD), переднее вентральное (n. AV), переднее медиальное (n. AM) и др. 2. Группа ядер средней линии включает в себя центральное медиальное (n. Cm), паравентрикулярное (n. Pv), ромбовидное (n. Rb) ядра, центральное серое вещество (Gc) и др. 3. Медиальная и интраламинарная группа содержит медиодорсальное (n. MD), центральное латеральное (n. CL), парацентральное (n. Pc) и другие ядра. 4. Вентролатеральная ядерная группа состоит из вентрального и латерального отделов. Вентральный отдел содержит вентральное переднее (n. VA), вентральное латеральное (n. VL) и вентральное заднее (n. VP) ядра. Латеральный отдел состоит из латерального дорсального (n. LD) и латерального заднего (n. LP) ядер. Здесь же находится ретикулярное ядро таламуса (n. R), ему принадлежит особое место в реализации функций таламуса. 5. Задняя группа ядер – подушечное ядро (PuCV), наружное и внутреннее коленчатые тела (n. GL, n. GM) и т.д. 6. Претектальная ядерная группа (иногда ее относят к задней группе ядер) содержит претектальное ядро (n. Prt), заднее ядро (n. P), претектальную зону и ядра задней спайки. С функциональной точки зрения все ядра таламуса подразделяют на 3 группы: 1 группа – специфические (релейные) ядра (сенсорные и несенсорные); 2 группа – неспецифические ядра; 3 группа – ассоциативные ядра. Специфические ядра имеют отчетливую топографическую и функциональную разграниченность проекций к определенным областям коры больших полушарий. Специфические ядра называют также релейными, переключающими. Специфические ядра делят на сенсорные релейные и несенсорные релейные. Несенсорные релейные ядра, в свою очередь, делятся на моторные ядра и переднюю группу. Отдельные морфологи переднюю группу и ряд неспецифических ядер называют лимбическими ядрами таламуса, учитывая их проекции на лимбическую кору. Например, специфические несенсорные ядра – переднее дорсальное, переднее медиальное и переднее вентральное – проецируются на различные поля поясной извилины. Релейные ядра таламуса получают афференты от лемнисковых систем (спинальной, тригемиальной, слуховой и зрительной), от некоторых структур головного мозга (вентральное переднее ядро таламуса, мозжечок, гипоталамус, полосатое тело) и имеют прямой выход в кору головного мозга (проекционные области, моторная и лимбическая кора). Каждое релейное ядро получает нисходящие волокна из собственной кортикальной проекционной зоны. Тем самым создается морфологическая основа для функциональных связей между таламическим ядром и его корковой проекцией в виде замкнутых нейронных кругов циркулирующего возбуждения, посредством которых осуществляются их взаимно регулирующие отношения. Нейронные поля релейных ядер таламуса содержат: 1) таламокортикальные релейные нейроны, аксоны которых идут в III и IV слоя коры; 2) длинноаксонные интегративные нейроны, аксоны которых дают коллатерали в ретикулярную формацию среднего мозга и другие ядра таламуса; 3) короткоаксонные нейроны, аксоны которых не выходят за пределы таламуса. Значительная часть нейронов релейных ядер отвечает только за стимуляцию определенной модальности, но имеются также и мультисенсорные нейроны. Релейным ядром для импульсации, несущей зрительную информацию, является наружное коленчатое тело, проецирующееся на зрительную кору (поля 17, 18, 19). Слуховые импульсы переключаются во внутреннем коленчатом теле. Проекционной корковой зоной являются поля 41, 42 и поперечная извилина Гешля. Вентральное переднее ядро таламуса (n. VA) получает обильную афферентацию из базальных ганглиев. Это ядро посылает прямые афференты к коре лобной области, оперкулуму и островку. Через это ядро проходят без переключения волокна от дорсомедиального ядра к лобной коре и к ретикулярному таламическому ядру. Благодаря вентральному переднему ядру хвостатое ядро проецируется на кору. Вентролатеральное ядро (n. VL) некоторыми авторами относится к одному из центров, который регулирует моторную активность и оказывает значительное влияние на активность пирамидных нейронов. Это ядро получает основные афференты по таламическому пучку лентикулярной петли, который начинается от нейронов внутреннего членика бледного шара. Другая часть афферентов приходит из красного и зубчатого ядер мозжечка. Из зубчатого ядра выходят прямые волокна, которые проходят красное ядро, а затем переключаются на нейроны рубро-таламического ядра и направляются в вентролатеральное ядро. Большое количество волокон к этому ядру приходит из ядра Кахала, расположенного в ретикулярной формации ствола мозга. |