Физиология центральной нервной системы
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
ЧАСТЬ 2 ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ В основе современных представлений о структуре и функции мозга лежит нейронная теория, которая рассматривает центральную нервную систему как совокупность синаптически связанных между собой нервных элементов – нейронов. Но, несмотря на то, что ЦНС представляет собой единое образование, сложность ее настолько велика, что для анализа ее функциональных свойств целесообразно разделение всего мозга на определенные области или отделы в соответствии с их микроструктурой, развитием, функцией и нейронной организацией. Такое деление в какой-то мере условно, так как нейроны, которые находятся в значительном отдалении друг от друга, могут быть более тесно связаны, чем нейроны, которые территориально близко расположены, в пределах одной и той же области. Например, клетки Беца двигательной коры имеют более тесные связи с мотонейронами спинного мозга, чем с нейронами других областей коры. Выяснение роли, специфических особенностей различных нервных образований, их взаимодействие является предметом изучения частной физиологии ЦНС. В центральной нервной системе выделяют следующие отделы (рис. 2.1): – спинной мозг – medulla spinalis; – задний мозг – metencephalon; – продолговатый мозг – medulla oblongets Bulbus medulla spinalis; – средний мозг – mesencephalon; – промежуточный мозг – diencephalon; – передний мозг – prosencephalon, telencehhalon. 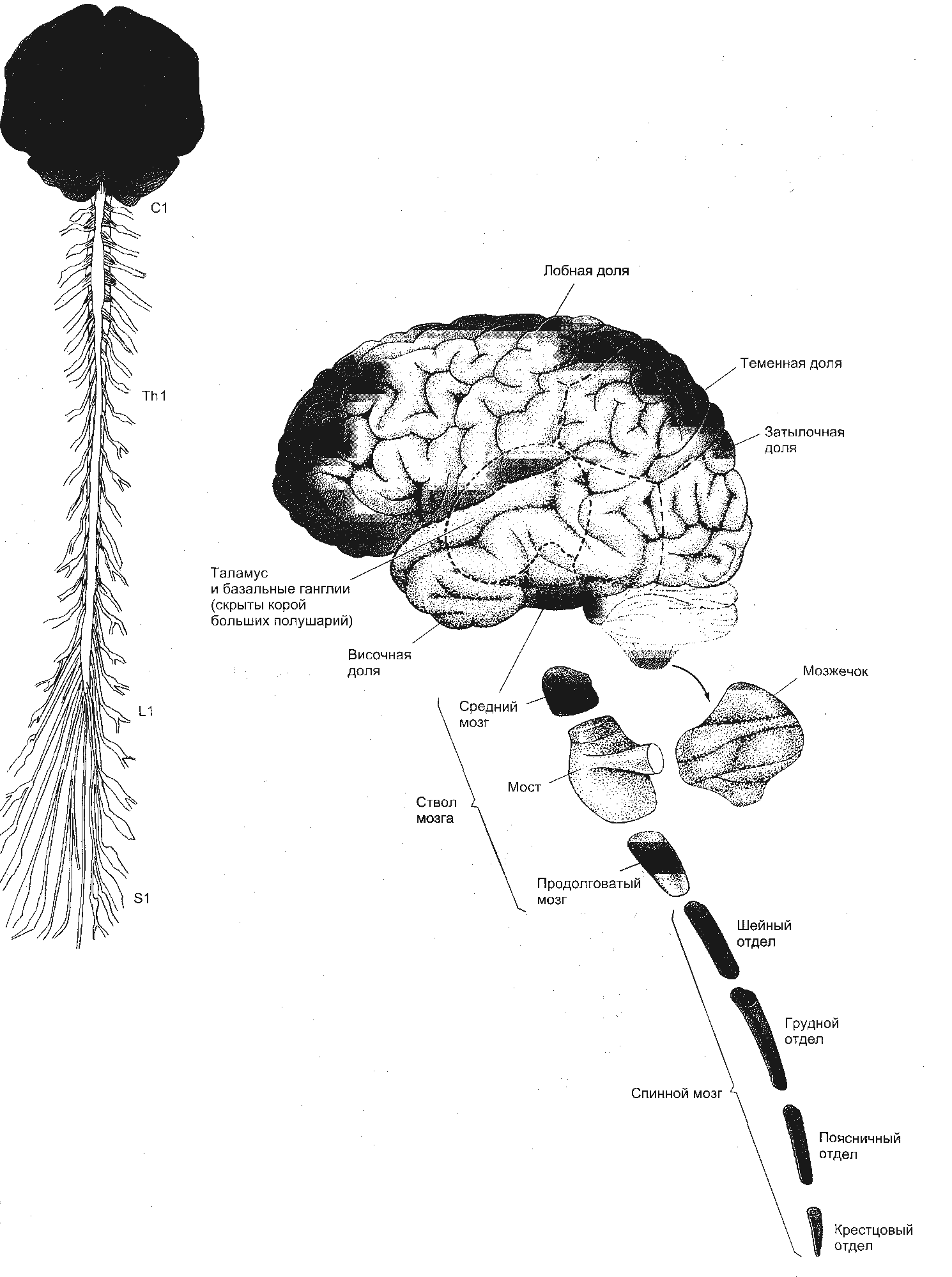 Рис. 2.1. Общий план организации центральной нервной системы и подразделение ее на отделы 7. Физиология спинного мозга 7.1. Морфофункциональная организация спинного мозга Спинной мозг – наиболее древний отдел ЦНС позвоночных. Он уже есть у ланцетника, наиболее примитивного представителя хордовых. Спинной мозг – каудальный отдел ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных. У человека в каудальном отделе позвоночного канала собираются корешки каудальных отделов спинного мозга, образуя так называемый конский хвост. Спинной мозг характеризуется сегментарным строением. В спинном мозге выделяют шейный, грудной, поясничный, крестцовый и копчиковый отделы. Каждый отдел состоит из нескольких сегментов. Шейный отдел включает 8 сегментов (С1 – С8), грудной – 12 (Th1 – Th12), поясничный – 5 (L1 – L5), крестцовый – 5 (S1 – S5) и копчиковый – 1-3 (Со1 – Co3). От каждого сегмента отходят две пары корешков, которые соответствуют одному из позвонков и покидают позвоночный канал через отверстие между ними. Различают дорсальные (задние) и вентральные (передние) корешки. Дорсальные корешки образованы центральными аксонами первичных афферентных нейронов, тела которых лежат в спинномозговых ганглиях. Вентральные корешки сформированы аксонами α- и γ-мотонейронов и немиелинизированными волокнами нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено независимо друг от друга в начале XIX века Ч. Беллом (1811) и Ф. Мажанди (1822). Различное распределение функций в передних и задних корешках спинного мозга получило название закона Белла-Мажанди. Сегменты спинного мозга и позвонки соответствуют одному метамеру. Нервные волокна пары задних корешков идут не только к своему метамеру, но также выше и ниже – в соседние метамеры. Кожная область, в которой распределяются эти чувствительные волокна, называется дерматомом. Количество волокон в дорсальном корешке гораздо больше, чем в вентральном. Нейронные структуры спинного мозга. Центральную часть поперечного среза спинного мозга занимает серое вещество. Вокруг серого вещества располагается белое вещество. В сером веществе выделяют передние, задние и боковые рога, а в белом – столбы (вентральные, дорсальные, латеральные и т.д.). Нейрональный состав спинного мозга достаточно разнообразен. Выделяют несколько видов нейронов. Тела нейронов спинномозговых ганглиев находятся вне спинного мозга. В спинной мозг входят аксоны этих нейронов. Нейроны спинномозговых ганглиев – это униполярные или псевдоуниполярные нейроны. В спинномозговых ганглиях лежат тела соматических афферентов, иннервирующих в основном скелетные мышцы. Тела других чувствительных нейронов находятся в ткани и в интрамуральных ганглиях автономной нервной системы и обеспечивают чувствительность только внутренних органов. Они бывают двух типов: крупные – диаметром 60-120 мкм и мелкие – диаметром 14-30 мкм. Крупные дают миелинизированные волокна, а мелкие – миелинизированные и немиелинизированные. Нервные волокна чувствительных клеток по скорости проведения и диаметру классифицируются на А-, В- и С-волокна. Толстые миелинизированные А-волокна диаметром от 3 до 22 мкм и скоростью проведения от 12 до 120 м/с подразделяются на подгруппы: альфа-волокна – от мышечных рецепторов, бета-волокна – от тактильных и барорецепторов, дельта-волокна – от терморецепторов, механорецепторов и болевых рецепторов. К волокнам группы В относят миелинизированные волокна средней толщины со скоростью проведения возбуждения 3-14 м/с. По ним в основном передается ощущение боли. К афферентам типа С относят большинство безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения до 2 м/с. Это волокна, которые идут от болевых, хемо- и некоторых механорецепторов. В сером веществе спинного мозга выделяют следующие элементы: 1) эфферентные нейроны (мотонейроны); 2) вставочные нейроны; 3) нейроны восходящих трактов; 4) интраспинальные волокна чувствительных афферентных нейронов. Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро тянется обычно на протяжении нескольких сегментов, поэтому и их аксоны, которые иннервируют одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков. В промежуточной зоне серого вещества локализуются вставочные нейроны. Их аксоны распространяются как внутри сегмента, так и в ближайшие соседние сегменты. Вставочные нейроны – разнородная группа, дендриты и аксоны которой не покидают пределов спинного мозга. Вставочные нейроны образуют синаптические контакты только с другими нейронами, и их большинство. На долю вставочных нейронов приходится около 97% от всех нейронов. По размерам они меньше α-мотонейронов, способны к высокочастотной импульсации (выше 1000 в сек.). Для проприоспинальных вставочных нейронов характерно свойство посылать длинные аксоны через несколько сегментов и оканчиваться на мотонейронах. Вместе с тем на эти клетки конвергируют волокна различных нисходящих трактов. Поэтому они являются релейными станциями на пути от вышележащих нейронов к мотонейронам. Особую группу вставочных нейронов образуют тормозные нейроны. К ним относятся, например, клетки Реншоу. Нейроны восходящих трактов также целиком находятся в пределах ЦНС. Тела этих нейронов расположены в сером веществе спинного мозга. Центральные окончания первичных афферентов имеют свои особенности. После вступления в спинной мозг афферентное волокно, как правило, дает начало восходящей и нисходящей ветвям, которые могут идти на значительные расстояния по спинному мозгу. Концевые разветвления одного нервного афферентного волокна имеют многочисленные синапсы на одном мотонейроне. Кроме того, установлено, что одно волокно, идущее от рецептора растяжения, образует синапсы почти со всеми мотонейронами данной мышцы. В дорсальной части дорсального рога располагается желатинозная субстанция Роланда. Наиболее точное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои или пластины, в каждой из которых группируются, как правило, однотипные нейроны. Согласно этим данным, все серое вещество спинного мозга было разделено на 10 пластин (Рексед) (рис. 2.2). I – краевые нейроны – дают начало спиноталамическому тракту; II-III – желатинозная субстанция; I-IV – в целом первичная сенсорная область спинного мозга (афферентация от экстерорецепторов, афферентация от рецепторов кожной и болевой чувствительности); 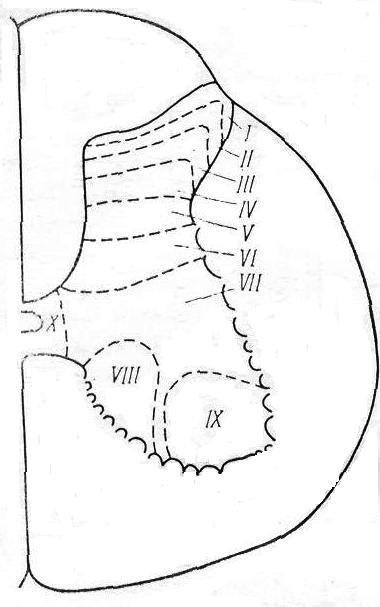 Рис. 2.2. Деление серого вещества спинного мозга на пластины (по Рекседу) V-VI – локализуются вставочные нейроны, которые получают входы от задних корешков и нисходящих путей (кортико-спинальный, рубро-спинальный); VII-VIII – располагаются проприоспинальные вставочные нейроны (от проприорецепторов, волокон вестибуло-спинального и ретикуло-спи- нального трактов), аксоны проприоспинальных нейронов; IX – содержит тела α- и γ-мотонейронов, пресинаптические волокна первичных афферентов от мышечных рецепторов растяжения, окончания волокон нисходящих трактов; X – окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон. Свойства нервных элементов спинного мозга. Спинной мозг человека содержит примерно 13 миллионов нейронов. α-мотонейроны – крупные клетки с длинными дендритами, имеющие до 20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Скорость проведения по их аксону составляет 70-120 м/с. Характерны ритмические разряды с частотой не выше 10-20 имп/с, что связано с выраженной следовой гиперполяризацией. Это выходные нейроны. Они осуществляют передачу сигналов скелетным мышечным волокнам, выработанных в спинном мозге. γ-мотонейроны – более мелкие клетки. Диаметр их не более 30-40 мкм, они не имеют непосредственного контакта с первичными афферентами. γ-мотонейроны иннервируют интрафузальные (внутриверетенные) мышечные волокна. Они моносинаптически активируются волокнами нисходящих трактов, что играет важную роль в α-, γ-взаимодействии. Скорость проведения по их аксону ниже – 10-40 м/с. Частота импульсов выше, чем у α-мото- нейронов, – 300-500 имп/с. В боковых и передних рогах находятся преганглионарные нейроны вегетативной нервной системы – аксоны их направляются к клеткам ганглиев симпатической нервной цепочки и к интрамуральным ганглиям внутренних органов. Тела симпатических нейронов, аксоны которых образуют преганглионарные волокна, располагаются в интермедиолатеральном ядре спинного мозга. Их аксоны относятся к группе В-волокон. Им свойственна постоянная тоническая импульсация. Одни из этих волокон участвуют в поддержании сосудистого тонуса, а другие обеспечивают регуляцию висцеральных эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых клеток). Тела парасимпатических нейронов образуют крестцовые парасимпатические ядра. Они располагаются в сером веществе крестцового отдела спинного мозга. Для многих из них характерна фоновая импульсная активность, частота которой возрастает, например, по мере повышения давления в мочевом пузыре. 7.2. Проводниковая функция спинного мозга Проводниковая функция спинного мозга осуществляется посредством проводящих путей, составляющих белое вещество спинного мозга. Проводящие пути подразделяются на следующие виды: восходящие и нисходящие тракты; тракты или пути, осуществляющие связь между нейронами спинного мозга, – проприоспинальные тракты. Важнейшие восходящие (сенсорные) тракты. Их больше, чем нисходящих (более 20), много еще неизученного, классификация уточняется. Эти пути проводят импульсы от рецепторов, воспринимающих информацию из внешнего мира и внутренней среды организма. Рассмотрим некоторые из них. 1. Тонкий пучок (пучок Голля) находится в задних столбах белого вещества спинного мозга. По этому пути проводится возбуждение от тактильных рецепторов, виброрецепторов, рецепторов положения тела в пространстве (проприоцептивный тракт). Этот тракт заканчивается в одноименном ядре продолговатого мозга. 2. Клиновидный пучок (пучок Бурдаха) имеет те же функции, что и тонкий пучок. Заканчивается клиновидный пучок в одноименном ядре продолговатого мозга. Короткие аксоны обоих пучков устанавливают синаптические связи с мотонейронами и интернейронами своего сегмента, а длинные аксоны направляются в продолговатый мозг. По пути они отдают большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя, таким образом, межсегментарные связи. По волокнам тонкого пучка проводится возбуждение от каудальной части тела и тазовых конечностей; по волокнам клиновидного пучка – от краниальной части тела и грудных конечностей. В спинном мозге оба эти пути проходят не прерываясь и не перекрещиваясь. В клиновидном и тонком ядрах продолговатого мозга происходит переключение на второй нейрон. Отростки второго нейрона направляются к специфическим ядрам таламуса противоположной стороны, образуя тем самым своеобразный перекрест. Здесь они переключаются на третий нейрон, аксоны которого достигают IV слоя коры больших полушарий. Считают, что по этой системе проводится информация тонкодифференцированной чувствительности, позволяющей определить локализацию и контур периферического раздражителя, а также его изменение во времени. 3. Дорсолатеральный спиноталамический тракт проходит в боковых столбах белого вещества спинного мозга. Это пути болевой и температурной чувствительности. 4. Вентральный спиноталамический тракт проходит в передних столбах белого вещества спинного мозга. Проводит возбуждение от тактильных рецепторов. Спиноталамические тракты либо прерываются и перекрещиваются на уровне сегмента, в который они только что вступили, либо вначале проходят несколько сегментов по своей стороне, а затем переходят на противоположную. Отсюда волокна идут к ядрам таламуса. В таламусе они образуют синапсы на нервных клетках, аксоны которых направляются в кору больших полушарий. Полагают, что по системе волокон этих путей в основном передается информация о качественной природе раздражителей. Существует мнение, что по спиноталамическим путям возможна передача возбуждения от проприо- и висцерорецепторов. Скорость проведения составляет 1-30 м/c. 5. Дорсальный спиномозжечковый (Кларка) тракт проходит в боковых столбах белого вещества спинного мозга. Данный тракт проводит импульсы от проприорецепторов мышц, сухожилий, связок, от рецепторов давления и прикосновения к коже. 6. Вентральный спиномозжечковый тракт, или пучок Говерса. Через продолговатый мозг и ножки мозжечка волокна этого тракта направляются к коре мозжечка, где занимают обширную площадь. Импульсы со скоростью до 120 м/с идут в кору мозжечка от сухожилий, от кожных и висцерорецепторов. Данные тракты участвуют в поддержании тонуса мышц и сохранения позы. Спиномозжечковых трактов более 10. Все они проводят импульсы от рецепторов, которые обеспечивают информацией высшие отделы центральной нервной системы для организации программ движения. К важнейшим нисходящим (моторным) трактам относятся следующие: 1. Пирамидный, или кортико-спинальный, тракт. Он является прямым, начинается от 5 слоя моторной коры, гигантских пирамид (клеток Беца) и идет к мотонейронам спинного мозга. Основная часть волокон перекрещивается на уровне пирамид продолговатого мозга. Остальные волокна перекрещиваются на уровне соответствующих сегментов спинного мозга. Моносинаптически оказывают облегчающий эффект на α-мотоней- роны мышц-сгибателей. 2. Рубро-спинальный тракт. Данный тракт берет начало от красного ядра (nucleus ruber) среднего мозга. Он полностью перекрещенный. Перекрест осуществляется сразу после выхода аксонов из красного ядра. Часть волокон идет в мозжечок и ретикулярную формацию ствола, а другие в спинной мозг. Моносинаптически рубро-спинальный тракт оказывает возбуждающий эффект на α-мотонейроны мышц-сгибателей. Рубро-спи- нальный тракт несет информацию от мозжечка, вестибулярных ядер и полосатого тела. 3. Вестибуло-спинальные тракты. Они берут начало от вестибулярных ядер продолговатого мозга (ядро Швальбе, ядро Бехтерева, ядро Дейтерса). Наибольшее значение имеет тракт, образованный ядром Дейтерса. Этот тракт также полностью перекрещенный. Перекрест начинается на уровне продолговатого мозга. Моносинаптически вестибуло-спиналь- ный тракт оказывает облегчающий эффект на α-мотонейроны мышц-разгибателей. 4. Ретикуло-спинальные тракты. Эти тракты начинаются от различных ядер ретикулярной формации продолговатого мозга, моста и среднего мозга. Ретикуло-спинальные тракты являются билатеральными, но гомолатеральные волокна доминируют. В ретикулярной формации продолговатого мозга имеются структуры, раздражение которых вызывает диффузный эффект – торможение α-мотонейронов сгибателей и разгибателей или их облегчение. Имеются структуры, которые функционируют реципрокно. От каудального ядра моста берет начало медиальный тракт; от латерального ядра – латеральный ретикуло-спинальный тракт, который перекрещивается в среднем мозге и спускается в спинной мозг. Медиальный ретикуло-спинальный тракт не перекрещивается. Он проецируется в вентральный столб. Латеральный тракт активирует α-мотонейроны, а медиальный – γ-мотонейроны. Раздражение различных зон ретикулярной формации ствола мозга вызывает как возбуждающие, так и тормозящие влияния в спинном мозге, как диффузные нереципрокные, так и реципрокные. Ретикуло-спинальные тракты, берущие начало в покрышке среднего мозга (например, тегменто-спинальный тракт), отдают коллатерали в ретикулярную формацию моста и продолговатого мозга, доходят до 7-10 грудных сегментов спинного мозга и нисходят в вентральных столбах спинного мозга. Данные тракты усиливают разряд γ-мотонейронов мышц-разгиба- телей. При раздражении покрышки среднего мозга, где берут начало эти тракты, возникает поза, характерная для ориентировочного рефлекса. |