мат мод хищник жертва. Исследование элементарной модели хищникжертва
 Скачать 0.88 Mb. Скачать 0.88 Mb.
|

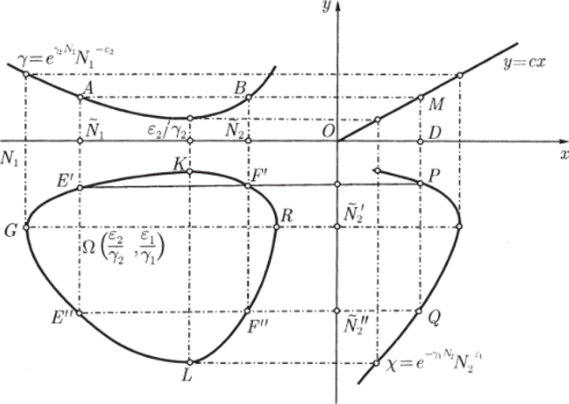 Рис. 3.3. через N до пересечения с кривой (3.3.10) (см. рис. 3.3). В свою очередь, значению К(Д^) соответствует некоторая точка М на прямой Y = сХ и, следовательно, некоторое значение X(N) = Y(N)/c, которое можно найти, проведя перпендикуляры AM и MD. Найденному значению (эта точка отмечена на рисунке буквой D) соответствуют две точки Р и G на кривой (3.3.11). По этим точкам, проводя перпендикуляры, найдем сразу две точки Е" и Е ", лежащие на кривой (3.3.9). Их координаты:  Проводя перпендикуляр AM , мы пересекли кривую (3.3.10) еще в одной точке В. Этой точке соответствуют те же Р и Q на кривой (3.3.11) и те же N и Щ. Координату N этой точки можно найти, опустив перпендикуляр из В на ось ON. Таким образом, мы получим точки F" и F", также лежащие на кривой (3.3.9). Исходя из другой точки N, тем же самым образом получим новую четверку точек, лежащих на кривой (3.3.9). Исключение составит точка Ni = ?2/72- Исходя из нее, получим только две точки: К и L. Это будут нижняя и верхняя точки кривой (3.3.9). Можно исходить не из значений N , а из значений N 2 . Направляясь от N 2 к кривой (3.3.11), поднимаясь затем до прямой У = сХ, а оттуда пересекая кривую (3.3.10), также найдем четыре точки кривой (3.3.9). Исключение составит точка No = ?1/71- Исходя из нее, получим только две точки: G и К. Это будут самая левая и самая правая точки кривой (3.3.9). Задавая разные N и N 2 и получив достаточно много точек, соединив их, приближенно построим кривую (3.3.9). Из построения видно, что эго замкнутая кривая, содержащая внутри себя точку 12 = (?2/721 ?1/71)» исходящая из определенных начальных данных N ю и N20. Взяв другое значение С, т.е. другие начальные данные, получим другую замкнутую кривую, не пересекающую первую и также содержащую точку (?2/721 ?1/71)1 внутри себя. Таким образом, семейство траекторий (3.3.9) есть семейство замкнутых линий, окружающих точку 12 (см. рис. 3.3). Исследуем тип устойчивости этой особой точки, воспользовавшись методом Ляпунова. Так как все параметры е 1, ?2, 71,72 положительны, точка (N[расположена в положительном квадранте фазовой плоскости. Линеаризация системы вблизи этой точки дает: 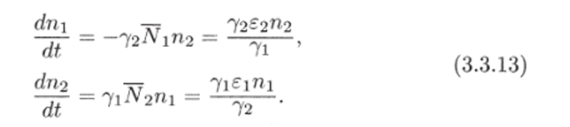 Здесь n(t) и 7i2(N1, N 2 : Характеристическое уравнение системы (3.3.13): 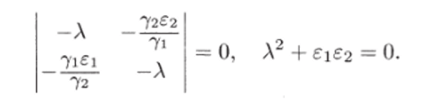 Корни этого уравнения чисто мнимые: Таким образом, исследование системы показывает, что траектории вблизи особой точки представлены концентрическими эллипсами, а сама особая точка - центр (рис. 3.4). Рассматриваемая модель Вольтерра и вдали от особой точки имеет замкнутые траектории, хотя форма этих траекторий уже отличается от эллипсоидальной. Поведение переменных Ni, N 2 во времени показано на рис. 3.5. 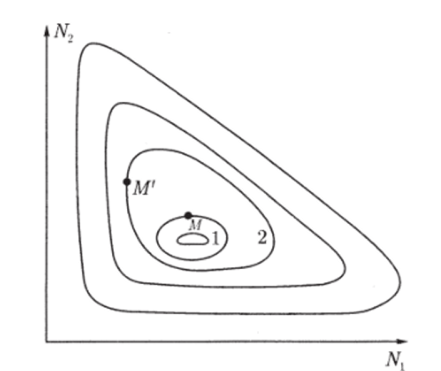 Рис. 3.4.  Рис. 3.5. Зависимость численности жертвы N i и хищника N 2 от времени Особая точка типа центр устойчива, но не асимптотически. Покажем на данном примере, в чем это заключается. Пусть колебания Ni(t) и ЛГгМ происходят таким образом, что изображающая точка движется по фазовой плоскости по траектории 1 (см. рис. 3.4). В момент, когда точка находится в положении М, в систему извне добавляется некоторое количество особей N 2, такое, что изображающая точка переходит скачком из точки М в точку Л/". После этого, если система снова предоставлена самой себе, колебания Ni и N 2 уже будут происходить с большими амплитудами, чем прежде, и изображающая точка двигается по траектории 2. Это и означает, что колебания в системе неустойчивы: они навсегда изменяют свои характеристики при внешнем воздействии. В дальнейшем мы рассмотрим модели, описывающие устойчивые колебательные режимы, и покажем, что на фазовой плоскости такие асимптотические устойчивые периодические движения изображаются при помощи предельных циклов. На рис. 3.6 изображены экспериментальные кривые - колебания численности пушных зверей в Канаде (по данным компании Гудзонова залива). Эти кривые построены на основании данных по числу заготовленных шкурок. Периоды колебаний численности зайцев (жертв) и рысей (хищников) примерно одинаковы и порядка 9 10 лет. При этом максимум численности зайцев опережает, как правило, максимум численности рысей на один год. Форма этих экспериментальных кривых значительно менее правильная, чем теоретических. Однако в данном случае достаточно того, что модель обеспечивает совпадение наиболее существенных характеристик теоретических и экспериментальных кривых, г.е. величин амплитуды и сдвига фаз между колебаниями численностей хищников и жертв. Гораздо более серьезным недостатком модели Вольтерра является неустойчивость решений системы уравнений. Действительно, как уже говорилось выше, любое случайное изменение численности того или другого вида должно привести, следуя модели, к изменению амплитуды колебаний обоих видов. Естественно, что в природных условиях животные подвергаются бесчисленному количеству таких случайных воздействий. Как видно из экспериментальных кривых, амплитуда колебаний численностей видов мало изменяется от года к году. Модель Вольтерра - эталонная (базовая) для математической экологии в той же мере, в какой модель гармонического осциллятора является базовой для классической и квантовой механики. 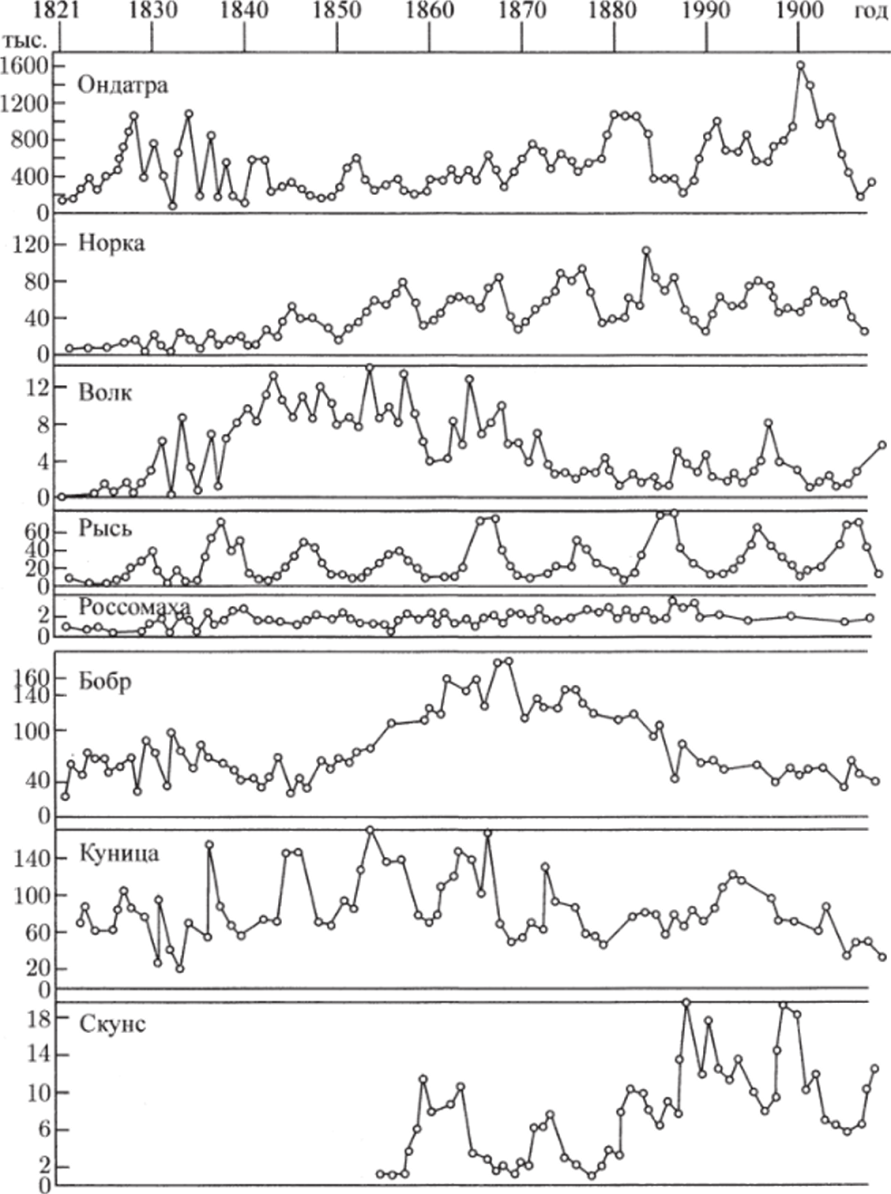 Рис. 3.6. Кинетические кривые численности пушных зверей по данным пуш ной компании Гудзонова залива (Сетон-Томсон, 1987) ческими средствами было выведено заключение о качественном характере поведения такой системы - о наличии в такой системе колебаний численности популяции. Без построения математической модели и ее использования такой вывод был бы невозможен. В рассмотренном нами выше самом простом виде системе Воль- терра присущи два принципиальных и взаимосвязанных недостатка. Их «устранению» посвящена обширная эколого-математическая литература. Во-первых, включение в модель любых, сколь угодно малых, дополнительных факторов качественным образом меняет поведение системы. Второй «биологический» недостаток модели заключается в том, что в нее не включены принципиальные свойства, присущие любой паре взаимодействующих по принципу хищник-жертва популяций: эффект насыщения хищника, ограниченность ресурсов хищника и жертвы даже при избытке жертвы, возможность минимальной численности жертв, доступных для хищника, и пр. С целью устранения этих недостатков были предложены разными авторами различные модификации системы Вольтерра. Наиболее ин- тересные из них будут рассмотрены в разделе 3.5. Здесь остановимся лишь на модели, учитывающей самоограничения в росте обеих популяций. На примере этой модели наглядно видно, как может меняться характер решений при изменении параметров системы. Итак, рассматривается система 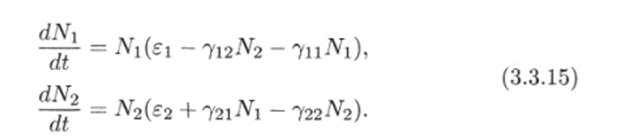 Система (3.3.15) отличается от ранее рассмотренной системы (3.3.6) наличием в правых частях уравнений членов вида -7uNf, Эти члены отражают тот факт, что численность популяции жертв не может расти до бесконечности даже в отсутствие хищников в силу ограниченности пищевых ресурсов, ограниченности ареала существования. Такие же «самоограничения» накладываются и на популяцию хищников. Для нахождения стационарных численностей видов iVi и N 2 приравняем к нулю правые части уравнений системы (3.3.15). Решения с нулевыми значениями численностей хищников или жертв не будут нас сейчас интересовать. Поэтому рассмотрим систему алгебраических Уравнений 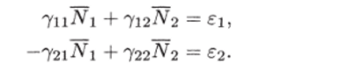 Ее решение дает нам координаты особой точки. На параметры системы здесь следует положить условие положительности стационарных численностей: N > 0 и N 2 > 0. Корни характеристического уравнения системы, линеаризованной в окрестности особой точки (3.3.16):  Из выражения для характеристических чисел видно, что если выполнено условие то численности хищников и жертв совершают во времени затухающие колебания, система имеет ненулевую особую точку устойчивый фокус. Фазовый портрет такой системы изображен на рис. 3.7 а. Допустим, что параметры в неравенстве (3.3.17) так изменяют свои значения, что условие (3.3.17) обращается в равенство. Тогда характеристические числа системы (3.3.15) равны, а ее особая точка будет лежать на границе между областями устойчивых фокусов и узлов. При изменении знака неравенства (3.3.17) на обратный особая точка становится устойчивым узлом. Фазовый портрет системы для этот случая представлен на рис. 3.76. Как и в случае одной популяции, для модели (3.3.6) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения JVj, N 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке 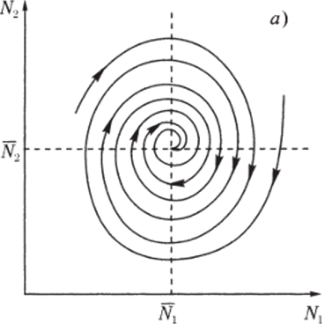 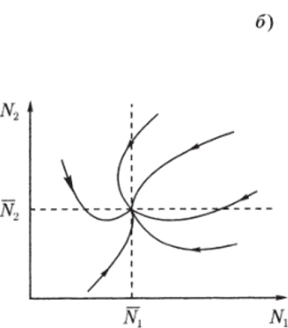 Рис. 3.7. Фазовый портрет системы (3.3.15): а - при выполнении соотношения (3.3.17) между параметрами; б - при выполнении обратного соотношения между параметрами фазовой траектории какая-либо переменная не очень велика, то существенное значение могут приобрести случайные флуктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический «дрейф» рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник- жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934; 2000), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum - хищника. Ожидавшиеся согласно детерминистическим уравнениям (3.3.6) равновесные численности в этих экспериментах составляли примерно всего но пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники). Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. В модели хищник жертва незатухающие колебания появляются вследствие выбора специальной формы уравнений модели (3.3.6). При этом модель становится негрубой, что свидетельствует об отсутствии в такой системе механизмов, стремящихся сохранить ее состояние. Однако в природе и эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. Рассмотрению таких обобщенных моделей посвящен раздел 3.5. Хищники могут поедать травоядных животных, и также слабых хищников. Хищники обладают широким спектром питания, легко переключаются с одной добычи на другую, более доступную. Хищники часто нападают на слабые жертвы. Поддерживается экологическое равновесие между популяциями жертва-хищник. Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает. Пресс хищников особенно силен, когда в коэволюции хищник - жертва равновесие смещается в сторону хищника и ареал жертвы сужается. Конкурентная борьба тесно связана с нехваткой пищевых ресурсов, она может быть и прямой борьбой, например, хищников за пространство как ресурс, но чаще всего это просто вытеснение вида, которому на данной территории пищи не хватает, видом, которому этого же количества пищи вполне достаточно. Это уже межвидовая конкуренция. И окончательно в системе ’’хищник - жертва”, описываемой моделью (2.7), возникновение диффузионной неустойчивости (при локальной устойчивости равновесия) возможно лишь в том случае, когда естественная смертность хищника возрастает с ростом его численности быстрее, чем линейная функция, и трофическая функция отличается от вольтерровской либо, когда популяция жертвы - это популяция типа Олли Теоретически в моделях «один хищник - две жертвы» эквивалентное выедание (отсутствие предпочтения того или иного вида жертвы) может повлиять на конкурентное сосуществование видов-жертв лишь в тех местах, где уже существует потенциально устойчивое равновесие . Разнообразие может возрасти только в таких условиях, когда у видов с меньшей конкурентоспособностью скорость роста популяции выше, чем у доминантов. Это позволяет понять ситуацию, когда равномерное выедание ведет к увеличению видового разнообразия растений там, где большее число видов, прошедших отбор на быстрое размножение, сосуществует с видами, эволюция которых направлена на повышение конкурентоспособности . Точно так же выбор жертвы, зависящий от ее плотности, может привести к устойчивому равновесию в теоретических моделях двух конкурирующих видов жертвы, где прежде никакого равновесия не существовало . Для этого хищник должен был бы обладать способностью к функциональным и численным реакциям на изменения плотности жертвы; возможно, однако, что переключение (непропорционально частые нападения на наиболее обильную жертву) будет иметь при этом более важное значение. В самом деле, установлено, что переключение оказывает стабилизирующее влияние в системах «один хищник - п жертв» и представляет собой единственный механизм, способный стабилизировать взаимодействия в тех случаях, когда ниши жертв полностью перекрываются . Такую роль могут играть неспециализированные хищники. Предпочтение более специализированными хищниками доминантного конкурента действует таким же образом, как переключение хищника, и может стабилизировать теоретические взаимодействия в моделях, в которых прежде не существовало равновесия между видами жертвы, при условии что их ниши в какой-то степени разделены . Также- не стабилизирует сообщество и хищник ’’сильный во всех отношениях”, т.е. хорошо адаптированный к данной жертве и с низкой относительной смертностью. При этом система обладает неустойчивым предельным циклом и, несмотря на устойчивость положения равновесия, вырождается в случайной среде (хищник выедает жертву и вследствие этого гибнет). Такая ситуация соответствует медленному вырождению. Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник-жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов. Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно «хищники» и «жертвы». Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности «хищников» приводит к уменьшению численности «жертв», это, в свою очередь, снижает запасы пищи («жертв») для «хищников», что в целом диктует снижение численности «жертв» и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды. Это в конечном итоге приходит к экологическому равновесию между популяциями хищника и жертвы. Для трофической функции третьего типа состояние равновесия будет устойчивым, если где N - точка перегиба функции (см. рис. 2, в). Это следует из того, что на интервале трофическая функция вогнута и, следовательно, относительная доля потребления хищником жертвы возрастает. Пусть Гг = -Г, т.е. имеет место сообщества типа ’’хищник - жертва”. В этом случае первое слагаемое в выражении (7.4) равно нулю и для выполнения условия устойчивости по вероятности равновесного состояния N требуется, чтобы и второе слагаемое не было положительным. Таким образом, для рассмотренного сообщества типа хищник - жертва можно сделать вывод об асимптотической устойчивости в целом положительного положения равновесия, т. е. для любых начальных данных 1Ч(0)>0 эволюция происходит таким образом, что N(7) - ■ К при условии, что N >0. Так, в однородной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. Волны жизни” (изменений численности хищника и жертвы) следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается примерно на одном уровне. Длительность периода зависит от скоростей роста обоих видов и от исходных параметров. Для популяции жертвы влияние хищника положительно, так как ее чрезмерное размножение привело бы к краху ее численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника. Другие модификации могут быть следствием особенностей поведения хищника. Число особей жертвы, которое хищник в состоянии потребить в данное время, имеет свой предел. Эффект насыщения хищника при приближении к этому рубежу показан в табл. 2-4, В. Взаимодействия, описываемые уравнениями 5 и 6, могут иметь устойчивые точки равновесия или обнаруживать циклические колебания. Однако такие циклы отличаются от тех, какие отражены в уравнениях Лотки - Вольтерры 1 и 2. Циклы, передаваемые уравнениями 5 и 6, могут иметь постоянную амплитуду и средние плотности, пока среда постоянна; после того как произошло нарушение, они могут вернуться к прежним амплитудам и средним плотностям. Такие циклы, которые восстанавливаются после нарушений, называются устойчивыми предельными циклами. Взаимодействие зайца и рыси можно считать устойчивым предельным циклом, но это не цикл Лотки - Вольтерры. Рассмотрим возникновение диффузионной неустойчивости в системе ’’хищник -жертва”, но сначапа выпишем условия, обеспечивающие возникновение диффузионной неустойчивости в системе (1.1) при п = 2. Ясно, что равновесие (N , Щ) локально Перейдем к интерпретации случаев, связанных с длительным сосуществованием хищника и жертвы. Понятно, что в отсутствие предельных циклов устойчивому равновесию будут соответствовать в случайной среде флуктуации численности, причем их амплитуда будет пропорциональна дисперсии возмущений. Такое явление будет происходить, если хищник имеет высокую относительную смертность и в то же время высокую степень приспособленности к данной жертве. Рассмотрим теперь, как меняется динамика системы с ростом приспособленности хищника, т.е. с убыванием Ъ от 1 до 0. Если приспособленность достаточно низкая, то предельные циклы отсутствуют, а равновесие является неустойчивым. С ростом приспособленности в окрестности этого равновесия возможно появление устойчивого цикла и далее внешнего неустойчивого. В зависимости от начальных условий (соотношения биомассы хищника и жертвы) система может либо терять устойчивость, т.е. уходить из окрестности равновесия, либо в ней будут со временем устанавливаться устойчивые колебания. Дальнейший рост приспособленности делает невозможным колебательный характер поведения системы. Однако при Ъ . Пример отрицательной (стабилизирующей) обратной связи - взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О -> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна. Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы. Если выполняется одно из условий: 0 1/2. Если же 6 > 1 . Устойчивость биоты и окружающей среды зависит только от взаимодействия растений - автотрофов и растительноядных гетеротрофных организмов. Хищники любых размеров не способны нарушить экологическое равновесие сообщества, так как в естественных условиях они не могут увеличить свою численность при постоянной численности жертв. Хищники не только должны быть сами передвигающимися, но и могут питаться только передвигающимися животными. Никакие другие рыбы не распространены так широко, как щуки. В немногих местах товли в стоячих или проточных водоемах нет давления со стороны щук для сохранения равновесия между жертвой и хищником Только современные искусственные водоемы, в которых щуки являются рыбами нежелательными из-за разведения там других рыб, не заселяются ими целенаправленно. В мире щуки исключительно хорошо представлены. Их ловят по всему северном) полушарию от Соединенных Штатов и Канады в |