мат мод хищник жертва. Исследование элементарной модели хищникжертва
 Скачать 0.88 Mb. Скачать 0.88 Mb.
|

|
МОДЕЛЬ «ХИЩНИК - ЖЕРТВА» И МАКРОЭКОНОМИЧЕСКАЯ МОДЕЛЬ ГУДВИНА Рассмотрим биологическую модель «хищник - жертва», в которой один вид является кормом для другого. Эта модель, давно ставшая классической, была построена в первой половине XX в. итальянским математиком В. Вольтерра для объяснения колебаний уловов рыбы в Адриатическом море . В модели предполагается, что число хищников растет до тех пор, пока у них будет достаточно пищи, а увеличение поголовья хищников приводит к уменьшению популяции рыб-жертв. Когда последних становится мало, численность хищников уменьшается. Вследствие этого с некоторого момента начинается рост численности рыб-жертв, который через некоторое время вызывает рост популяции хищников. Цикл замыкается. Пусть N x (t) и N 2 (t) - численности рыб-жертв и рыб-хищников в момент времени t соответственно. Предположим, что темп прироста численности жертв в условиях отсутствия хищников постоянен, т. е. где а - положительная постоянная. Появление хищника должно снизить темп прироста жертв. Будем считать, что это снижение линейно зависит от численности хищника: чем больше хищников, тем меньше темп прироста жертв. Тогда где т > 0. Поэтому для динамики численности рыб-жертв получаем: Составим теперь уравнение, определяющее динамику популяции хищников. Предположим, что их численность в случае отсутствия жертв сокращается (из-за отсутствия корма) с постоянным темпом Ь, т. е. Наличие жертв вызывает увеличение темпа прироста хищников. Предположим, что этот прирост линеен, т. е. где п> 0. Тогда для скорости роста рыб-хищников получаем уравнение: В системе «хищник - жертва» (6.17)-(6.18) снижение скорости роста численности рыб-жертв, вызванное поеданием их хищниками, равно mN x N 2 , т. е. пропорционально числу их встреч с хищником. Увеличение скорости роста численности рыб-хищников, вызванное наличием жертв, равно nN x N 2 , т. е. тоже пропорционально числу встреч жертв и хищников. Введем безразмерные переменные U = mN 2 /a и V = nN x /b. Динамика переменной U соответствует динамике хищников, а динамика переменной V - динамике жертв. В силу уравнений (6.17) и (6.18) изменение новых переменных определяется системой уравнений: Предположим, что при t = 0 число особей обоих видов известно, следовательно, известны начальные значения новых переменных?/(0) = U 0 , К(0) = К 0 . Из системы уравнений (6.19) можно найти дифференциальное уравнение для ее фазовых траекторий: 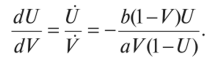 Разделяя переменные этого уравнения, получим:  Рис. 6.10. Построение фазовой траектории ADCBA системы дифференциальных уравнений (6.19) Отсюда с учетом начальных данных следует: где константа интегрирования С = b(V Q - InV 0)/a - lnU 0 + U 0 . На рис. 6.10 показано, как строится линия (6.20) при заданном значении С. Для этого в первой, во второй и в третьей четвертях соответственно строим графики функций х = V - InV, у = (Ь/а)х , у = = InU- U+C. В силу равенства dx/dV = (V- 1)/У функция х = V- In К, определенная при V > 0, возрастает, если V> 1, и убывает, если V 1. В силу того, что cPx/dV 1 = 1/F 2 > 0, график функции л: = x(V) направлен выпуклостью вниз. Уравнение V= 0 задает вертикальную асимптоту. Наклонных асимптот эта функция не имеет. Поэтому график функции х = х(У) имеет вид кривой, изображенной в первой четверти рис. 6.10. Аналогичным образом исследуется функция у = InU - U+ С, график которой на рис. 6.10 изображен в третьей четверти. Если теперь мы разместим на рис. 6.10 во второй четверти график функции у = (Ь/а)х , то в четвертой четверти получим линию, которая связывает переменные U и V. Действительно, взяв точку V t на оси OV, вычисляем с помощью функции х = V - V соответствующее знание х х. После этого, используя функцию у = (Ь/а)х , по полученному значению х { находим у х (вторая четверть на рис. 6.10). Далее с помощью графика функции у = InU - U + С определяем соответствующие значения переменной U (на рис. 6.10 таких значений два - координаты точек М и N). Совокупность всех таких точек (V; U) образует искомую кривую. Из построения следует, что графиком зависимости (6.19) является замкнутая линия, содержащая внутри себя точку Е( 1, 1). Напомним, что эту кривую мы получили, задав некоторые начальные значения U 0 и V 0 и вычислив по ним константу С. Взяв другие начальные значения, мы получим другую замкнутую линию, не пересекающую первую и также содержащую внутри себя точку Е( 1, 1). Это означает, что семейством траекторий системы (6.19) на фазовой плоскости (V, U) является множество замкнутых непересекающихся линий, концентрирующихся вокруг точки Е( 1, 1), а решения исходной модели U = Щ) и V = V{t) являются функциями, периодическими во времени. При этом максимум функции U = U(t) не попадает на максимум функции V = V(t ) и наоборот, т. е. колебания численности популяций около своих равновесных решений происходят в разных фазах. На рис. 6.11 приведены четыре траектории системы дифференциальных уравнений (6.19) на фазовой плоскости OUV, отличающиеся начальными условиями. Одна из траекторий равновесная - это точка Е( 1, 1), которой соответствует решение U(t) = 1, V{t) = 1. Точки (U(t), V(t)) на других трех фазовых траекториях по мере увеличения времени смещаются по часовой стрелке. Чтобы пояснить действие механизма изменения численности двух популяций, рассмотрим траекторию ABCDA на рис. 6.11. Как видим, на участке АВ и хищников, и жертв мало. Поэтому здесь популяция хищников сокращается из-за недостатка корма, а популяция жертв растет. На участке ВС численность жертв достигает высоких значений, что приводит к росту численности хищников. На участке СА хищников много, и это влечет сокращение численности жертв. При этом после прохождения точки D численность жертв уменьшается на столько, что численность популяции начинает уменьшаться. Цикл замыкается. Модель «хищник - жертва» - пример структурно неустойчивой модели. Здесь малое изменение правой части одного из уравнений может привести к принципиальному изменению ее фазового портрета. 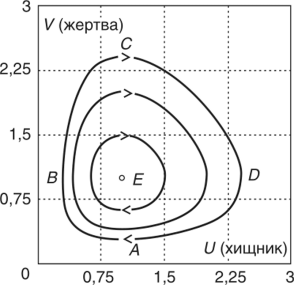 Рис. 6.11. 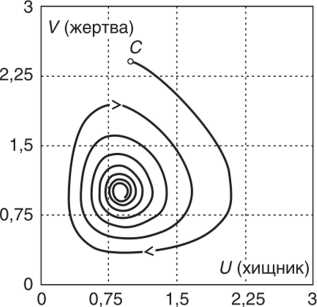 Рис. 6.12. Действительно, если в уравнении динамики жертв учесть внутривидовую конкуренцию, то мы получим систему дифференциальных уравнений: Здесь при т = 0 популяция жертв развивается согласно логическому закону. При т Ф 0 ненулевое равновесное решение системы (6.21) при некоторых положительных значениях параметра внутривидовой конкуренции И является устойчивым фокусом, и соответствующие траектории «наматываются» на точку равновесия (рис. 6.12). Если же h = 0, то в этом случае особая точка Е( 1, 1) системы (6.19) является центром, и траектории представляют собой замкнутые линии (см. рис. 6.11). Замечание. Обычно под моделью «хищник - жертва» понимают модель (6.19), фазовые траектории которой замкнуты. Однако модель (6.21) также является моделью «хищник - жертва», поскольку описывает взаимовлияние хищников и жертв. Одним из первых приложений модели типа «хищник - жертва» в экономике для исследования циклично изменяющихся процессов является макроэкономическая модель Гудвина, в которой используется непрерывный подход к анализу взаимовлияния уровня занятости и ставки заработной платы. В работе В.-Б. Занга изложен вариант модели Гудвина, в котором производительность труда и предложение труда растут с постоянными темпами прироста, а коэффициент выбытия фондов равен нулю. Эта модель формально приводит к уравнениям модели «хищник - жертва». Ниже рассматривается модификация этой модели для случая отличного от нуля коэффициента выбытия фондов. В модели используются следующие обозначения: L - численность трудящихся; w - средняя ставка заработной платы трудящихся; К - основные производственные фонды (капитал); Y - национальный доход; / - инвестиции; С - потребление; р - коэффициент выбытия фондов; N - предложение труда на рынке рабочей силы; т = Y/K - фондоотдача; а = Y/L - производительность труда; у = L/N - коэффициент занятости; х = C/Y - норма потребления в национальном доходе; К - приращение капитала в зависимости от инвестиций. Выпишем уравнения модели Гудвина: 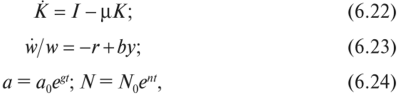 где а 0 , b, g, п, N 0 , г - положительные числа (параметры). Уравнения (6.22) - (6.24) выражают следующее. Уравнение (6.22) - обычное уравнение динамики фондов. Уравнение (6.23) отражает рост ставки заработной платы при высоком уровне занятости (ставка заработной платы растет, если предложение труда невелико) и уменьшение ставки заработной платы при высоком уровне безработицы. Тем самым уравнение (6.23) выражает закон Филлипса в линейной форме . Уравнения (6.24) означают экспоненциальный рост производительности труда и предложения труда. Предположим также, что С = wL, т. е. вся заработная плата тратится на потребление. Теперь можно преобразовать уравнения модели с учетом равенств: 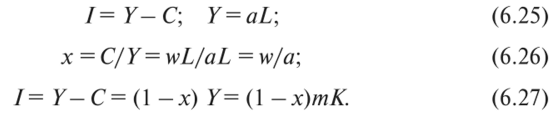 Преобразуем уравнения (6.22)-(6.27). Имеем: 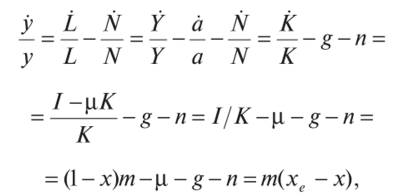 где Поэтому динамика переменных в модели Гудвина описывается системой дифференциальных уравнений:  которая формально совпадает с уравнениями классической модели «хищник - жертва». Это значит, что в модели Гудвина тоже возникают колебания фазовых переменных. Механизм колебательной динамики заключается здесь в следующем: при малой заработной плате w потребление низкое, инвестиции большие, и это приводит к росту производства и занятости у. Большая занятость у вызывает рост средней заработной платы w, что приводит к росту потребления и снижению инвестиций, падению производства и снижению занятости у. Ниже гипотеза о зависимости ставки процента от уровня занятости рассмотренной модели используется при моделировании динамики однопродуктовой фирмы. Оказывается, что в этом случае при некоторых дополнительных допущениях модель фирмы обладает свойством цикличности рассмотренной выше модели «хищник - жертва». Здесь в отличие от (3.2.1) знаки (-012) и (+a2i) разные. Как и в случае конкуренции (система уравнений (2.2.1)), начало координат (1) для этой системы является особой точкой типа «неустойчивый узел». Три других возможных стационарных состояния: 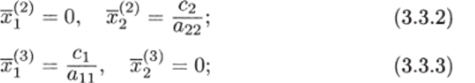 Биологический смысл требует положительности величин Х у х 2. Для выражения (3.3.4) это означает, что В случае, если коэффициент внутривидовой конкуренции хищников а ,22 = 0, условие (3.3.5) приводит к условию ai2 Возможные типы фазовых портретов для системы уравнений (3.3.1) представлены на рис. 3.2 a-в. Изоклины горизонтальных касательных представляют собой прямые а изоклины вертикальных касательных - прямые Из рис. 3.2 видно следующее. Система хищник -жертва (3.3.1) может иметь устойчивое положение равновесия, в котором популяция жертв полностью вымерла (х = 0) и остались только хищники (точка 2 на рис. 3.26). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х хищник Х 2 имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для хз. Особые точки (1) и (3) (рис. 3.26) являются неустойчивыми. Вторая возможность - устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы - устойчивая точка (3) (рис. 3.2а). Здесь особая точка (1) - также неустойчивый узел. Наконец, третья возможность - устойчивое сосуществование популяций хищника и жертвы (рис. 3.2 в), стационарные численности которых выражаются формулами (3.3.4). Рассмотрим этот случай подробнее. Предположим равенство нулю коэффициентов внутривидовой конкуренции (аи = 0, i = 1, 2). Предположим также, что хищники питаются только жертвами вида х и в отсутствие их вымирают со скоростью С2 (в (3.3.5) С2 Проведем подробное исследование этой модели, воспользовавшись обозначениями, наиболее широко принятыми в литературе. Переобо- 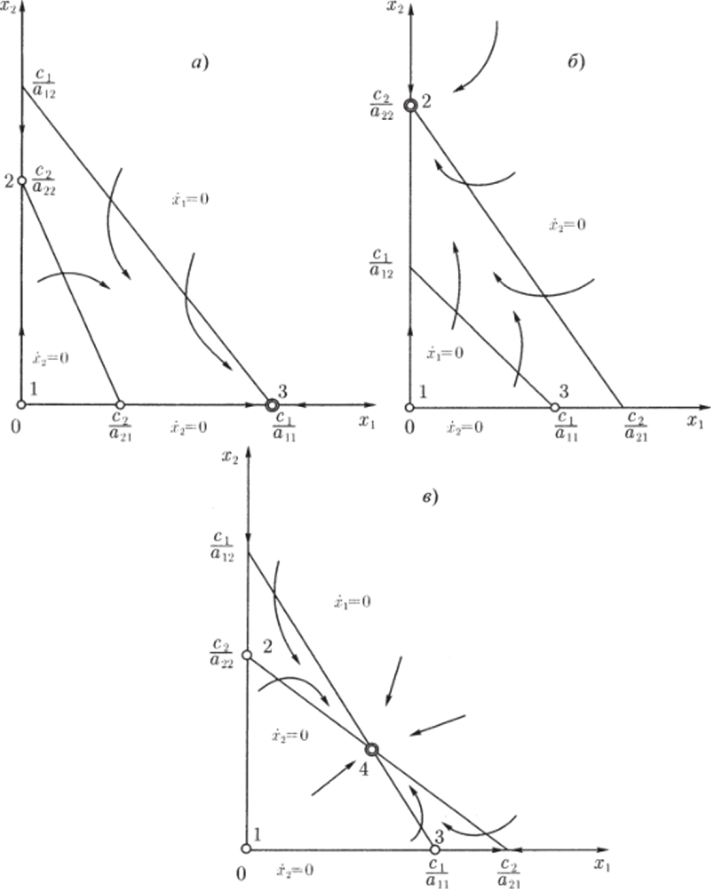 Рис. 3.2. Расположение главных изоклин на фазовом портрете вольтерров- ской системы хищник жертва при разном соотношении параметров: а - о» б - С I С2 С2 1, 3 - неустойчивые, 2 - устойчивая особая точка; в - 1, 2, 3 - неустойчивые, 4 - устойчивая особая точка значим Система хищник-жертва в этих обозначениях имеет вид: 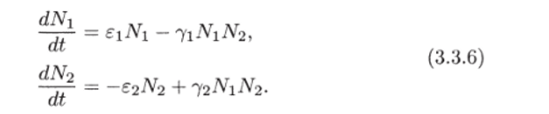 Свойства решений системы (3.3.6) будем исследовать на фазовой плоскости N 1 ON 2 Система имеет два стационарных решения. Их легко определить, приравняв нулю правые части системы. Получим: 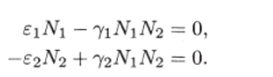 Отсюда стационарные решения: 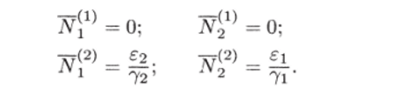 Рассмотрим подробнее второе решение. Найдем первый интеграл системы (3.3.6), не содержащий t. Умножим первое уравнение на -72, второе - на -71 и результаты сложим. Получим: Теперь разделим первое уравнение на N и умножим на € 2, а второе разделим на JV 2 и умножим на е. Результаты снова сложим: Сравнивая (3.3.7) и (3.3.8), будем иметь: 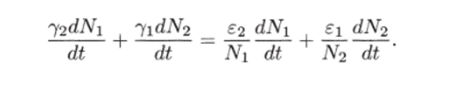 Интегрируя, получим:  Это и есть искомый первый интеграл. Таким образом, система (3.3.6) является консервативной, поскольку имеет первый интеграл движения, величину, представляющую собой функцию переменных системы N и N 2 и не зависящую от времени. Это свойство позволяет конструировать для вольтерровских систем систему понятий, аналогичную статистической механике (см. гл. 5), где существенную роль играет величина энергии системы, неизменная во времени. При каждом фиксированном с > 0 (что соответствует определенным начальным данным) интегралу соответствует определенная траектория на плоскости N 1 ON 2 , служащая траекторией системы (3.3.6). Рассмотрим графический способ построения траектории, предложенный самим Вольтерра. Заметим, что правая часть формулы (3.3.9) зависит только от Д г 2, а левая - только от N. Обозначим Из (3.3.9) следует, что между X и Y имеется пропорциональная зависимость На рис. 3.3 изображены первые квадранты четырех систем координат XOY, NOY , N 2 OX и Д Г 10N 2 так, чтобы все они имели общее начало координат. В левом верхнем углу (квадрант NOY) построен график функции (3.3.8), в правом нижнем (квадрант N 2 OX) - график функции Y. Первая функция имеет min при Ni = а вторая - max при N 2 = ?- Наконец, в квадранте XOY построим прямую (3.3.12) для некоторого фиксированного С. Отметим точку N на оси ON . Этой точке соответствует определенное значение Y(N 1), которое легко найти, проведя перпендикуляр |