мат мод хищник жертва. Исследование элементарной модели хищникжертва
 Скачать 0.88 Mb. Скачать 0.88 Mb.
|

4.5 Система ХИЩНИК+ЖЕРТВА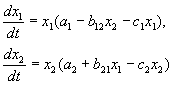 (9.7) (9.7)Здесь, в отличие от (9.2) знаки b 12 и b 21 - разные. Как и в случае конкуренции, начало координат является особой точкой типа неустойчивый узел. Три других возможных стационарных состояния: Таким образом, возможно выживание только жертвы (9.10), только хищника (9.9) (если у него имеются и другие источники питания) и сосуществование обоих видов (9.11). Последний вариант уже был рассмотрен нами в лекции 5. Возможные типы фазовых портретов для системы хищник-жертва представлены на рис. 9.4. Изоклины горизонтальных касательных представляют собой прямые x 2 = – b 21 х 1 /c 2 + a 1 /c 2 , х 2 = 0, а изоклины вертикальных касательных – прямые x 2 = – c 1 х 1 /b 12 + a 2 /b 12 , х 1 = 0. Стационарные точки лежат на пересечении изоклин вертикальных и горизонтальных касательных.  Из рис. 9.4 видно следующее. Система хищник – жертва (9.7) может иметь устойчивое положение равновесия, в кот o ром популяция жертв полностью вымерла ( ) и остались только хищники (точка 2 на рис. 9.4 а). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х 1 хищник х 2 – имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для х 2 . Особые точки (1) и (3) (рис. 9.4 а ) являются неустойчивыми. Вторая возможность – устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы – устойчивая точка (3) (рис. 9.4 6 ). Здесь особая точка (1) – также неустойчивый узел. Наконец, третья возможность – устойчивое сосуществование популяций хищника и жертвы (рис. 9.4 в ), стационарные численности которых выражаются формулами (9.11). Как и в случае одной популяции (см. Лекция 3), для модели (9.7) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения x 1 , x 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке фазовой траектории какая–либо переменная не очень велика, то существенное значение могут приобрести случайные флюктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический “дрейф” рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник – жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum – хищника. Ожидавшиеся согласно детерминистическим уравнениям (9.7) равновесные численности в этих экспериментах составляли примерно всего по пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники) Результаты экспериментов представлены на рис. 9.5. 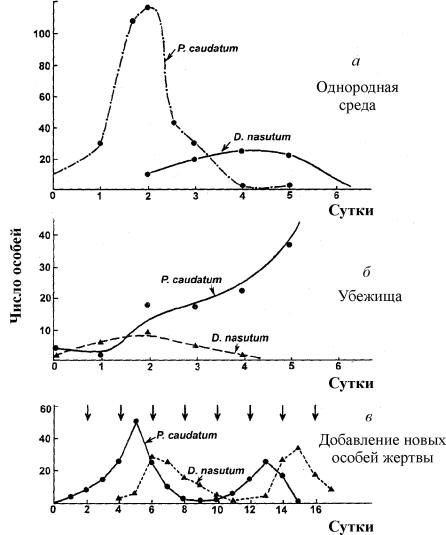 Рис . 9.5. Рост Parametium caudatum и хищной инфузории Dadinium nasutum . Из : Gause G.F. The struggle for existence. Baltimore , 1934 Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. Однако в природе и в эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. 4.6 Обобщенные модели взаимодействия двух видов Было предложено большое число моделей, описывающих взаимодействие видов, правые части уравнений которых представляли собой функции численностей взаимодействующих популяций. Решался вопрос о выработке общих критериев, позволяющих установить, какого вида функции могут описать особенности поведения временного численности популяции, в том числе устойчивые колебания. Наиболее известные из этих моделей принадлежат Колмогорову (1935, переработанная статья ‑ 1972) и Розенцвейгу (1963). 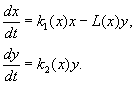 (9.12) (9.12)4.7 Модель Колмогорова В модель заложены следующие предположения: 1) Хищники не взаимодействуют друг с другом, т.е. коэффициент размножения хищников k 2 и число жертв L , истребляемых в единицу времени одним хищником, не зависит от y. 2) Прирост числа жертв при наличии хищников равен приросту в отсутствие хищников минус число жертв, истребляемых хищниками. Функции k 1 (x ), k 2 (x ), L (x ), ‑ непрерывны и определены на положительной полуоси x , y ³ 0. 3) dk 1 /dx< 0. Это означает, что коэффициент размножения жертв в отсутствие хищника монотонно убывает с возрастанием численности жертв, что отражает ограниченность пищевых и иных ресурсов. 4) dk 2 / dx> 0, k 2 (0) < 0 < k 2 (¥ ). С ростом численности жертв коэффициент размножения хищников монотонно убывает с возрастанием численности жертв, переходя от отрицательных значений, (когда нечего есть) к положительным. 5) Число жертв, истребляемых одним хищником в единицу времени L (x )> 0 при N> 0; L (0)=0. Возможные типы фазовых портретов системы (9.12) представлены на рис. 9.6:  Рис. 9.6. Фазовые портреты системы Колмогорова (9.12), описывающей взаимодействие двух видов при разных соотношениях параметров. Пояснения в тексте. Стационарные решения (их два или три) имеют следующие координаты: (1). ` x=0; ` y=0 . Начало координат при любых значениях параметров представляет собой седло(рис. 9.6 а-г). (2). ` x=A, ` y=0. (9.13) A определяется из уравнения: k 1 (A )=0. Стационарное решение (9.13) ‑ седло, если B< A (рис. 9.6 а , б , г ), B определяется из уравнения k 2 (B )=0 Точка (9.13) помещается в положительном квадранте, еслиB>A. Это ‑ устойчивый узел. Последний случай, который соответствует гибели хищника и выживанию жертвы,представлен на рис. 9.6 в . (3). ` x=B, ` y=C. (9.14) Величина С определяется из уравнений: Точка (9.14) – фокус (рис.9.6 а ) или узел (рис.9.6 г ), устойчивость которых зависит от знакавеличины s s 2 = – k 1 (B) – k 1 (B )B+L (B )C. Если s >0, точка устойчива, если s <0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 б ) В зарубежной литературе чаще рассматривается сходная модель, предложенная Розенцвейгом и Макартуром (1963): 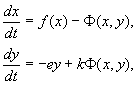 (9.15) (9.15)где f (x ) - скорость изменения численности жертв x в отсутствие хищников, Ф(x,y ) ‑ интенсивность хищничества, k ‑ коэффициент, характеризующий эффективность переработки биомассы жертвы в биомассу хищника, e - смертность хищника. Модель (9.15) сводится к частному случаю модели Колмогорова (9.12) при следующих предположениях: 1) численность хищника ограничивается только численностью жертвы, 2) скорость, с которой данная особь хищника поедает жертву, зависит только от плотности популяции жертв и не зависит от плотности популяции хищников. Тогда уравнения (9.15) принимают вид. 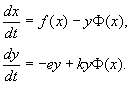 При описании взаимодействия реальных видов, правые части уравнений конкретизируются в соответствии с представлениями о биологических реалиях. Рассмотрим одну из самых популярных моделей такого типа. 4.8 Модель взаимодействия двух видов насекомых Макартура (M acArthur, 1971) Модель, которую мы рассмотрим ниже, использовалась для решения практической задачи борьбы с вредными насекомыми с помощью стерилизации самцов одного из видов. Исходя из биологических особенностей взаимодействия видов, была написана следующая модель 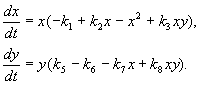 (9.16) (9.16)Здесь x,y - биомассы двух видов насекомых. Трофические взаимодействия описываемых в этой модели видов – весьма сложные. Этим определяется вид многочленов в правых частях уравнений. Рассмотрим правую часть первого уравнения. Насекомые вида х поедают личинок вида у (член + k 3 y), но взрослые особи вида у поедают личинок вида х при условии высокой численности видов х или у или обоих видов (члены – k 4 xy, – у 2 ). При малых х смертность вида х выше, чем его естественный прирост (1–k 1 +k 2 x–x 2 < 0 при малых х). Во втором уравнении член k 5 отражает естественный прирост вида y; –k 6 y – самоограничение этого вида, –k 7 x – поедание личинок вида у насекомыми вида х, k 8 xy – прирост биомассы вида у за счет поедания взрослыми насекомыми вида у личинок вида х. На рис. 9.7 представлен предельный цикл, являющийся траекторией устойчивого периодического решения системы (9.16). 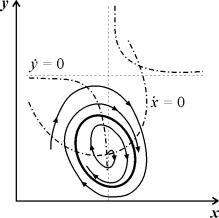 Решение вопроса о том, каким образом обеспечить сосуществование популяции с ее биологическим окружением, разумеется, не может быть получено без учета специфики конкретной биологической системы и анализа всех ее взаимосвязей. Вместе с тем изучение формальных математических моделей позволяет ответить на некоторые общие вопросы. Можно утверждать, что для моделей типа (9.12) факт совместимости или несовместимости популяций не зависит от их начальной численности, а определяется только характером взаимодействия видов. Модель помогает ответить на вопрос: как следует воздействовать на биоценоз, управлять им, чтобы по возможности быстро уничтожить вредный вид. Управление может сводиться к кратковременному, скачкообразному изменению величин численности х и у. Такой способ отвечает методам борьбы типа однократного уничтожения одной или обеих популяций химическими средствами. Из сформулированного выше утверждения видно, что для совместимых популяций этот метод борьбы будет малоэффективным, поскольку с течением времени система опять выйдет на стационарный режим. Другой способ ‑ изменение вида функций взаимодействия между видами, например, при изменении значений параметров системы. Именно такому, параметрическому, способу отвечают биологические методы борьбы. Так при внедрении стерилизованных самцов уменьшается коэффициент естественного прироста популяции. Если при этом мы получим другой тип фазового портрета, такой, где имеется лишь устойчивое стационарное состояние с нулевой численностью вредителя, управление приведет к желаемому результату – уничтожению популяции вредного вида. Интересно отметить, что иногда воздействие целесообразно применить не к самому вредителю, а к его партнеру. Какой из способов более эффективен, в общем случае сказать нельзя. Это зависит от имеющихся в распоряжении средств управления и от явного вида функций, описывающих взаимодействие популяций. 4.9 Модель А.Д.Базыкина Теоретический анализ моделей взаимодействий видов наиболее исчерпывающе проведен в книге А.Д.Базыкина “Биофизика взаимодействующих популяций” (М., Наука, 1985). Рассмотрим одну из изученных в этой книге моделей типа хищник-жертва. 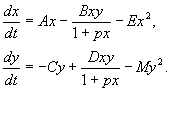 (9.17) (9.17)Система (9.17) является обобщением простейшей модели хищник-жертва Вольтерра (5.17) с учетом эффекта насыщения хищников. В модели (5.17) предполагается, что интенсивность выедания жертв линейно растет с ростом плотности жертв, что при больших плотностях жертв не соответствует реальности. Для описания зависимости рациона хищника от плотности жертв могут быть выбраны разные функции. Наиболее существенно, чтобы выбранная функция с ростом x асимптотически стремилась к постоянному значению. В модели (9.6) использовалась логистическая зависимость. В модели Базыкина в роли такой функции выбрана гипербола x /(1+px ). Вспомним, что такой вид имеет формула Моно, описывающая зависимость скорость роста микроорганизмов от концентрации субстрата. Здесь в роли субстрата выступает жертва, а в роли микроорганизмов – хищник. Система (9.17) зависит от семи параметров. Число параметров можно уменьшить с помощью замены переменных: x ® (A/D )x; y ® (A/D )/y; t ® (1/A )t; g (9.18) и зависит от четырех параметров. Для полного качественного исследования необходимо разбить четырехмерное пространство параметров на области с различным типом динамического поведения, т.е. построить параметрический, или структурный портрет системы. Затем надо построить фазовые портреты для каждой из областей параметрического портрета и описать бифуркации, происходящие с фазовыми портретами на границах различных областей параметрического портрета. Построение полного параметрического портрета производится в виде набора “срезов” (проекций) параметрического портрета малой размерности при фиксированных значениях некоторых из параметров. Параметрический портрет системы (9.18) при фиксированных g и малых e представлен на рис.9.8. Портрет содержит 10 областей с различным типом поведения фазовых траекторий.  Рис. 9.8. Параметрический портрет системы (9.18) при фиксированных g и малых e Поведение системы при различных соотношениях параметров может быть существенно различным (рис.9.9). В системе возможны: 1) одно устойчивое равновесие (области 1 и 5); 2) один устойчивый предельный цикл (области 3 и 8); 3) два устойчивых равновесия (область 2) 4) устойчивый предельный цикл и неустойчивое равновесие внутри него (области 6, 7, 9, 10) 5) устойчивый предельный цикл и устойчивое равновесие вне его (область 4). В параметрических областях 7, 9, 10 область притяжения равновесия ограничивается неустойчивым предельным циклом, лежащим внутри устойчивого. Наиболее интересно устроен фазовый портрет, соответствующий области 6 на параметрическом портрете. Детально он изображен на рис. 9.10.  Область притяжения равновесия В 2 (заштрихована) представляет собой “улитку”, скручивающуюся с неустойчивого фокуса В 1 . Если известно, что в начальный момент времени система находилась в окрестности В 1 , то судить о том, придет ли соответствующая траектория в равновесие В 2 или на устойчивый предельный цикл, окружающий три точки равновесия С (седло), В 1 и В 2 можно лишь на основе вероятностных соображений.  Рис.9.10. Фазовый портрет системы 9.18 для параметрической области 6. Область притяжения В 2 заштрихована На параметрическом портрете (9.7) имеются 22 различные бифуркационные границы, которые образуют 7 различных типов бифуркаций. Их изучение позволяет выявить возможные типы поведения системы при изменении ее параметров. Например, при переходе из области 1 в область 3 происходит рождение малого предельного цикла, или мягкое рождение автоколебаний вокруг единственного равновесия В. Аналогичное мягкое рождение автоколебаний, но вокруг одного из равновесий, а именно B 1 , происходит при пересечении границы областей 2 и 4. При переходе из области 4 в область 5 устойчивый предельный цикл вокруг точки B 1 “лопается” на петле сепаратрис и единственной притягивающей точкой остается равновесие B 2 и т.д. Особый интерес для практики представляет, конечно, выработка критериев близости системы к бифуркационным границам. Действительно, биологам хорошо известно свойство “буферности”, или “гибкости”, природных экологических систем. Этими терминами обычно обозначают способность системы как бы поглощать внешние воздействия. Пока интенсивность внешнего воздействия не превышает некоторой критической величины, поведение системы не претерпевает качественных изменений. На фазовой плоскости это соответствует возвращению системы в устойчивое состояние равновесия или на устойчивый предельный цикл, параметры которого не сильно отличаются от первоначального. Когда же интенсивность воздействия превышает допустимую, система “ломается”, переходит в качественно иной режим динамического поведения, например просто вымирает. Это явление соответствует бифуркационному переходу. Каждый тип бифуркационных переходов имеет свои отличительные особенности, позволяющие судить об опасности такого перехода для экосистемы. Приведем некоторые общие критерии, свидетельствующие о близости опасной границы. Как и в случае одного вида, если при уменьшении численности одного из видов происходит “застревание” системы вблизи неустойчивой седловой точки, что выражается в очень медленном восстановлении численности к начальному значению, значит, система находится вблизи критической границы. Индикатором опасности служит также изменение формы колебаний численностей хищника и жертвы. Если из близких к гармоническим колебания становятся релаксационными, причем амплитуда колебаний увеличивается, это может привести к потере устойчивости системы и вымиранию одного из видов. Дальнейшее углубление математической теории взаимодействия видов идет по линии детализации структуры самих популяций и учета временных и пространственных факторов. |