Лекция фрш. лекции ФРШ. Кафедра биологии лекционный комплекс по дисциплине физиологическое развитие школьников
 Скачать 1.66 Mb. Скачать 1.66 Mb.
|

|
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков: аксон пирамидного нейрона идет через белое вещество в другие зоны коры или структуры ЦНС. Звездчатые нейроны имеют короткие, хорошо ветвящиеся дендриты и короткий аксон, обеспечивающий связи нейронов в пределах самой коры большого мозга. Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры. Строение коры больших полушарий В коре содержится большое количество глиальных клеток, выполняющих опорную, обменную, секреторную, трофическую функции. Наружная поверхность коры разделена на четыре доли: лобную, теменную, затылочную и височную. Каждая доля имеет свои проекционные и ассоциативные области. Кора большого мозга имеет шестислойное строение (рис. 1): молекулярный слой (1) светлый, состоит из нервных волокон и имеет небольшое количество нервных клеток; наружный зернистый слой (2) состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре головного мозга, т.е. имеющих отношение к памяти; слой пирамидных меток (3) формируется из пирамидных клеток малой величины и вместе со слоем 2 обеспечивает корко-корко- вые связи различных извилин мозга; внутренний зернистый слой (4) состоит из звездчатых клеток, здесь заканчиваются специфические таламокортикальные пути, т.е. пути, начинающиеся от рецепторов-анализаторов. внутренний пирамидный слой (5) состоит из гигантских пирамидных клеток, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг; слой полиморфных клеток (6) состоит из неоднородных по величине клеток треугольной и веретенообразной формы, которые образуют кортикоталамические пути. 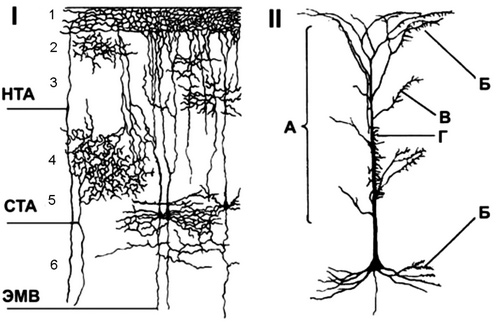 I — афферентные пути из таламуса: СТА — специфические таламические афференты; НТА — неспецифические таламические афференты; ЭМВ — эфферентные моторные волокна. Цифрами обозначены слои коры; II — пирамидный нейрон и распределение окончаний на нем: А — неспецифические афферентные волокна из ретикулярной формации и таламуса; Б — возвратные коллатерали от аксонов пирамидных нейронов; В — комиссуральные волокна из зеркальных клеток противоположного полушария; Г — специфические афферентные волокна из сенсорных ядер таламуса Рис. 1. Связи коры больших полушарий. Клеточный состав коры по разнообразию морфологии, функций, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны. Это позволило выделить в мозге человека 53 цитоархитектонических поля. Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе. Функциональной единицей коры является вертикальная колонка диаметром около 500 мкм. Колонка — зона распределения разветвлений одного восходящего (афферентного) таламокортикального волокна. Каждая колонка содержит до 1000 нейронных ансамблей. Возбуждение одной колонки тормозит соседние колонки.  Восходящий путь проходит через все корковые слои (специфический путь). Неспецифический путь также проходит через все корковые слои. Белое вещество полушарий расположено между корой и базальными ганглиями. Оно состоит из большого количества волокон, идущих в разных направлениях. Это проводящие пути конечного мозга. Различают три вида путей. проекционный — связывает кору с промежуточным мозгом и другими отделами ЦНС. Это восходящие и нисходящие пути; комиссуральный — его волокна входят в состав мозговых комиссур, которые соединяют соответствующие участки левого и правого полушарий. Входят в состав мозолистого тела; ассоциативный — связывает участки коры одного и того же полушария. Зоны коры больших полушарий По особенностям клеточного состава поверхность коры подразделяют на структурные единицы следующего порядка: зоны, области, подобласти, поля. Зоны коры головного мозга разделяются на первичные, вторичные и третичные проекционные зоны. В них расположены специализированные нервные клетки, к которым поступают импульсы от определенных рецепторов (слуховых, зрительных и т.д.). Вторичные зоны представляют собой периферические отделы ядер анализаторов. Третичные зоны получают обработанную информацию от первичных и вторичных зон коры больших полушарий и играют важную роль в регуляции условных рефлексов. В сером веществе коры больших полушарий различают сенсорные, моторные и ассоциативные зоны: сенсорные зоны коры больших полушарий — участки коры, в которых располагаются центральные отделы анализаторов: зрительная зона — затылочная доля коры больших полушарий; слуховая зона — височная доля коры больших полушарий; зона вкусовых ощущений — теменная доля коры больших полушарий; зона обонятельных ощущений — гиппокамп и височная доля коры больших полушарий. Соматосенсорная зона находится в задней центральной извилине, сюда приходят нервные импульсы от проприорецепторов мышц, сухожилий, суставов и импульсы от температурных, тактильных и других рецепторов кожи; моторные зоны коры больших полушарии — участки коры, при раздражении которых появляются двигательные реакции. Располагаются в передней центральной извилине. При ее поражении наблюдаются значительные нарушения движения. Пути, по которым импульсы идут от больших полушарий к мышцам, образуют перекрест, поэтому при раздражении моторной зоны правой стороны коры возникает сокращение мышц левой стороны тела; ассоциативные зоны — отделы коры, находящиеся рядом с сенсорными зонами. Нервные импульсы, поступающие в сенсорные зоны, приводят к возбуждению ассоциативных зон. Особенностью их является то, что возбуждение может возникать при поступлении импульсов от различных рецепторов. Разрушение ассоциативных зон приводит к серьезным нарушениям обучения и памяти. Речевая функция связана с сенсорными и двигательными зонами. Двигательный центр речи (центр Брока) находится в нижней части левой лобной доли, при его разрушении нарушается речевая артикуляция; при этом больной понимает речь, но сам говорить не может. Слуховой центр речи (центр Вернике) расположен в левой височной доле коры больших полушарий, при его разрушении наступает словесная глухота: больной может говорить, излагать устно свои мысли, но не понимает чужой речи; слух сохранен, но больной не узнает слов, нарушается письменная речь. Речевые функции, связанные с письменной речью — чтение, письмо, — регулируются зрительным центром речи, расположенным на границе теменной, височной и затылочной долей коры головного мозга. Его поражение приводит к невозможности чтения и письма. В височной доле находится центр, отвечающий за запоминание слое. Больной с поражением этого участка не помнит названия предметов, ему необходимо подсказывать нужные слова. Забыв название предмета, больной помнит его назначение, свойства, поэтому долго описывает их качества, рассказывает, что делают с этим предметом, но назвать его не может. Например, вместо слова «галстук» больной говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости». Функции лобной доли: управление врожденными поведенческими реакциями при помощи накопленного опыта; согласование внешних и внутренних мотиваций поведения; разработка стратегии поведения и программы действия; мыслительные особенности личности. Состав коры больших полушарий Кора больших полушарий головного мозга является высшей структурой ЦНС и состоит из нервных клеток, их отростков и нейроглии. В составе коры имеются звездчатые, веретенообразные и пирамидные нейроны. Благодаря наличию складок кора имеет большую поверхность. Выделяют древнюю кору (архикортекс) и новую кору (неокортекс). Кора состоит из шести слоев (рис. 2).  Рис. 2. Кора больших полушарий головного мозга Верхний молекулярный слой образован в основном дендритами пирамидных клеток нижележащих слоев и аксонами неспецифических ядер таламуса. На этих дендритах формируют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса. Наружный гранулярный слой образован мелкими звездчатыми клетками и частично малыми пирамидными клетками. Волокна клеток этого слоя расположены преимущественно вдоль поверхности коры, формируя кортикокортикальные связи. Слой пирамидных клеток малой величины. Внутренний гранулярный слой, образованный звездчатыми клетками. В нем заканчиваются афферентные таламокортикальные волокна, начинающиеся от рецепторов анализаторов. Внутренний пирамидный слой состоит из крупных пирамидных клеток, участвующих в регуляции сложных форм движения. Мультиформный слой состоит из верстеновидных клеток, образующих кортикоталамические пути. По функциональной значимости нейроны коры подразделяют на сенсорные, воспринимающие афферентные импульсы от ядер таламуса и рецепторов сенсорных систем; моторные, посылающие импульсы к подкорковым ядрам, промежуточному, среднему, продолговатому мозгу, мозжечку, ретикулярной формации и спинному мозгу; и промежуточные, осуществляющие связь между нейронами коры больших полушарий. Нейроны коры больших полушарий находятся в состоянии постоянного возбуждения, не исчезающего и во время сна. В кору больших полушарий, к сенсорным нейронам поступают импульсы от всех рецепторов организма через ядра таламуса. И каждый орган имеет свою проекцию или корковое представительство, расположенное в определенных областях больших полушарий. В коре больших полушарий имеется четыре чувствительные и четыре двигательные области. Нейроны двигательной коры получают афферентную импульсацию через таламус от мышечных, суставных и кожных рецепторов. Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути. У животных наиболее развита лобная область коры и ее нейроны участвуют в обеспечении целенаправленного поведения. Если удалить эту долю коры, животное становится вялым, сонливым. В височной области локализуется участок слуховой рецепции, и сюда поступают нервные импульсы от рецепторов улитки внутреннего уха. Область зрительной рецепции находится в затылочных долях коры головного мозга. Теменная область, внеядерная зона, играет важную роль в организации сложных форм высшей нервной деятельности. Здесь расположены рассеянные элементы зрительного и кожного анализаторов, осуществляется межанализаторный синтез. Рядом с проекционными зонами располагаются ассоциативные зоны, которые осуществляют связь между сенсорной и двигательной зонами. Ассоциативная кора принимает участие в конвергенции различных сенсорных возбуждений, позволяющей осуществлять сложную обработку информации о внешней и внутренней среде. ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ. Впервые предположение о рефлекторном характере деятельности высших отделов мозга было высказано И. М. Сеченовым, что позволило распространить рефлекторный принцип работы и на психическую деятельность человека. Идеи И. М. Сеченова получили экспериментальное подтверждение в трудах И. П. Павлова, который разработал метод объективной оценки функций высших отделов мозга — метод условных рефлексов. Высшая нервная деятельность — это совокупность безусловных и условных рефлексов, а также высших психических функций, которые обеспечивают адекватное поведение организма в изменяющихся природных и социальных условиях. Термин «высшая нервная деятельность» впервые введён в науку И. П. Павловым, считавшим его эквивалентным понятию «психическая деятельность». Все формы психической активности, включая мышление и сознание человека, Павлов считал элементами высшей нервной деятельности. Основная роль в осуществлении высшей нервной деятельности у высших животных и человека принадлежит коре больших полушарий. Экспериментально показано, что у высших представителей животного мира после полного оперативного удаления коры высшая нервная деятельность резко ухудшается. Они теряют способность тонко приспосабливаться к внешней среде и самостоятельно существовать в ней. Чем выше уровень организации животного, тем большее влияние на его жизнедеятельность оказывает кора больших полушарий. Человек, лишённый коры больших полушарий (анэнцефалия), практически нежизнеспособен. Высшая нервная деятельность включает в себя совокупность условных и безусловных рефлексов, мышление, память, логику, речь и эмоции. Высшая нервная деятельность. Это деятельность высших отделов центральной нервной системы, обеспечивающая наиболее совершенное приспособление животных и человека к окружающей среде. Структурную основу высшей нервной деятельности у человека составляет кора больших полушарий вместе с ближайшими к ней подкорковыми образованиями. Функции этих отделов – осуществление сложных рефлекторных реакций. Безусловные рефлексы относительно постоянны, проявляются в ответ на адекватное раздражение и служат основой для формирования многочисленных условных рефлексов. Безусловные рефлексы обеспечивают координированную деятельность, направленную на поддержание внутренней среды и взаимодействие организма с внешней средой. Безусловные рефлексы – это врожденные, наследственно передающиеся реакции организма. Прежде всего, они делятся на простые и сложные. Особой группой были выделены сложнейшие безусловные рефлексы к числу которых были отнесены индивидуальные (пищевой, оборонительный, исследовательский и др.) и видовые (половой и родительский). Особое место среди безусловных рефлексов занимает ориентировочный рефлекс или рефлекс на новизну. Он возникает в ответ на любое достаточно быстро происходящее изменение окружающей среды и выражается внешне в настораживании, прислушивании, повороте глаз и головы, а иногда и всего тела в сторону появившегося нового раздражителя и т.п. Отличием ориентировочного рефлекса от других безусловно рефлекторных реакций является то, что он сравнительно быстро ослабевает - угасает при повторных – применениях одного и того же раздражителя. Таким образом, безусловный рефлекс - это реакция организма на непосредственно действующие раздражители, способствующая взаимодействию организма с окружающей средой и имеющая адаптивное значение для него. Условные рефлексы. Условные рефлексы - реакции, приобретенные организмом в процессе индивидуального развития на основе «жизненного опыта». Носят приспособительный характер, что делает поведение наиболее пластичным, приспособленным к конкретным условиям среды. Любые условные рефлексы требуют для своего участия высших отделов головного мозга, приобретаются и отменяются в индивидуальной жизни каждой конкретной особи, имеют сигнальный характер, т.е. предупреждают последующее возникновение безусловного рефлекса, подготавливая к нему организм. Условные рефлексы, различающиеся по особенностям безусловногоДля выработки условного рефлекса необходимы условия: условный раздражитель должен предшествовать безусловному, значимость безусловного раздражителя должна быть больше условного, нормальное функционирование головного мозга. подкрепления. В зависимости от наличия или отсутствия подкрепления условные рефлексы делятся на положительные (подкрепляемые), вызывающие соответствующую реакцию организма, и отрицательные или тормозные (неподкрепляемые), которые не только не вызывают соответствующей реакции, но и ослабляют ее. Если условный рефлекс выработан на базе безусловного, то он является рефлексом первого порядка. Если же на базе ранее выработанного условного рефлекса, то его именуют условным рефлексом второго порядка. Соответственно возможны условные рефлексы более высоких порядков. У детей описаны рефлексы 6-го порядка. У взрослых людей формируются условные рефлексы 2-20-го порядка. Различия безусловных и условных рефлексов. Безусловные рефлексы являются видовыми, т.е. свойственными всем представителям данного вида. Условные рефлексы индивидуальны: у одних представителей одного и того же вида они могут быть, а у других отсутствуют. Безусловные рефлексы относительно постоянны; условные непостоянны и в зависимости от определенных условий могут вырабатываться, закрепиться или исчезнуть. Осуществляются безусловные рефлексы в ответ на адекватные раздражения, приложенные к определенному рецептивному полю. Условные рефлексы могут образовываться на любые воспринимаемые организмом раздражения любого рецептивного поля. Условные рефлексы вырабатываются на базе безусловных. Они являются преимущественно функцией коры большого мозга. Безусловные рефлексы могут осуществляться на уровне спинного мозга и мозгового ствола. Возрастные особенности условных рефлексов. Условные рефлексы в период новорожденности носят очень ограниченный характер. Уже в первые дни жизни ребенка можно отметить образование натурального условного рефлекса на время кормления, выражающееся в пробуждении детей и повышенной двигательной активности. При строгом режиме кормления на 6-7 день у младенцев происходит условно-рефлекторное повышение количества лейкоцитов уже за 30 минут до кормления, и повышается газообмен перед приемом пищи. С середины первого месяца жизни возникают условные рефлексы на различные первосигнальные стимулы: свет, звук, обонятельные раздражения. Скорость образования условных рефлексов на первом месяце жизни очень мала и быстро увеличивается с возрастом. У детей дошкольного возраста значительно возрастает роль подражательного и игрового рефлекса. Так играя в куклы, дети точно копируют жесты, слова, манеры, воспитателей, родителей и т.д. Скорость образования условных рефлексов у детей старше 10 лет и у взрослых практически не отличается. В подростковом периоде затрудняется образование временных связей, а также уменьшается скорость образования условных рефлексов. Поэтому особенности высшей нервной деятельности требует внимательного к ним отношения, продуманной организации учебно-воспитательного процесса. Взаимодействие процессов возбуждения и торможения в центральной нервной системе обеспечивает точность и гибкость высшей нервной деятельности. По своему внешнему проявлению торможение противоположно возбуждению. Различают безусловное и условное торможение. Внешнее торможение. Безусловное торможение называют внешним или индукционным и оно свойственно всем элементам нервной системы и является врожденным. Этот вид торможения представляет собой процесс экстренного ослабления или прекращения отдельных поведенческих реакций при действии раздражителей, поступающих из внешней или внутренней среды. Внешнее торможение часто встречается в условиях повседневной жизни человека. Это постоянно наблюдаемое снижение активности, нерешительность действий в новой, необычной обстановке, снижение эффекта той или иной деятельности при действии каких-либо необычных посторонних раздражителей. Например, болевое раздражение или сигнал о нем резко тормозит пищевые условные рефлексы. В школьной практике условные рефлексы детей, связанные, например, с письмом, тормозятся, если на учащихся действует какой-либо достаточно сильный посторонний раздражитель. Таким раздражителем может быть, например, удар грома, громкий окрик учителя, чувство голода, духота и т. п. Внешнее торможение включает в себя гаснущий и постоянный тормоз, а так же запредельное торможение. |