|
|
Раздел 2. Клетка как биологическая система
Включения
Включения — это непостоянные компоненты клетки, которые образу�ются и исчезают в процессе ее жизнедеятельности. К ним относят как за�пасные вещества, например, зерна крахмала (рис. 2.41) или белка в рас�тительных клетках, гранулы гликогена в клетках животных и грибов, волютина у бактерий, капли жира во всех типах клеток, так и отходы жизнедеятельности, в частности, непереваренные в результате фагоцито�за остатки пищи, образующие так называемые остаточные тельца.
Взаимосвязь строения и функций частей и органоидов клетки — основа ее целостности
Каждая из частей клетки, с одной стороны, является обособленной структурой со специфи�ческим строением и функциями, а с другой — компонентом более сложной системы, называе�мой клеткой. Большая часть наследственной информации эукариотической клетки сосредоточена в ядре, однако само ядро не в состоянии обеспечить ее реализацию, поскольку для этого необхо�димы как минимум цитоплазма, выступающая как основное вещество, и рибосомы, на которых и происходит этот синтез. Большинство рибосом расположено на гранулярной эндоплазматической сети, откуда белки чаще всего транспортируются в комплекс Гольджи, а затем после моди�фикации — в те части клетки, для которых они предназначены. Мембранные упаковки белков и углеводов могут встраиваться в мембраны органоидов и цитоплазматическую мембрану, обеспе�чивая их постоянное обновление. От комплекса Гольджи отшнуровываются также выполняющие важнейшие функции лизосомы и вакуоли. Например, без лизосом клетки быстро превратились бы в своеобразную свалку отработанных молекул и структур.
Протекание всех этих процессов требует энергии, вырабатываемой митохондриями, а у расте�ний — и хлоропластами. И хотя эти органоиды являются относительно автономными, т. к. имеют собственные молекулы ДНК, часть их белков все равно кодируется ядерным геномом и синтези�руется в цитоплазме.
Таким образом, клетка представляет собой неразрывное единство составляющих ее компонен�тов, каждый из которых выполняет свою уникальную функцию.
2.5. Метаболизм: энергетический и пластический обмен, их взаимосвязь. Ферменты, их химическая природа, роль в метаболизме. Стадии энергетического обмена. Брожение и дыхание. Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле.
2.5.1. Энергетический и пластический обмен, их взаимосвязь.
2.5.2. Энергетический обмен в клетке (диссимиляция).
2.5.3. Фотосинтез и хемосинтез.
Обмен веществ и превращения энергии — свойства живых организмов
Клетку можно уподобить миниатюрной химической фабрике, на которой происходят сотни и тысячи химических реакций.
Обмен веществ — совокупность химических превращений, направленных на сохранение и са�мовоспроизведение биологических систем.
Он включает в себя поступление веществ в организм в процессе питания и дыхания, внутри�клеточный обмен веществ, или метаболизм, а также выделение конечных продуктов обмена.
Обмен веществ неразрывно связан с процессами превращения одних видов энергии в другие. Например, в процессе фотосинтеза световая энергия запасается в виде энергии химических свя�зей сложных органических молекул, а в процессе дыхания она высвобождается и расходуется на синтез новых молекул, механическую и осмотическую работу, рассеивается в виде тепла и т. д.
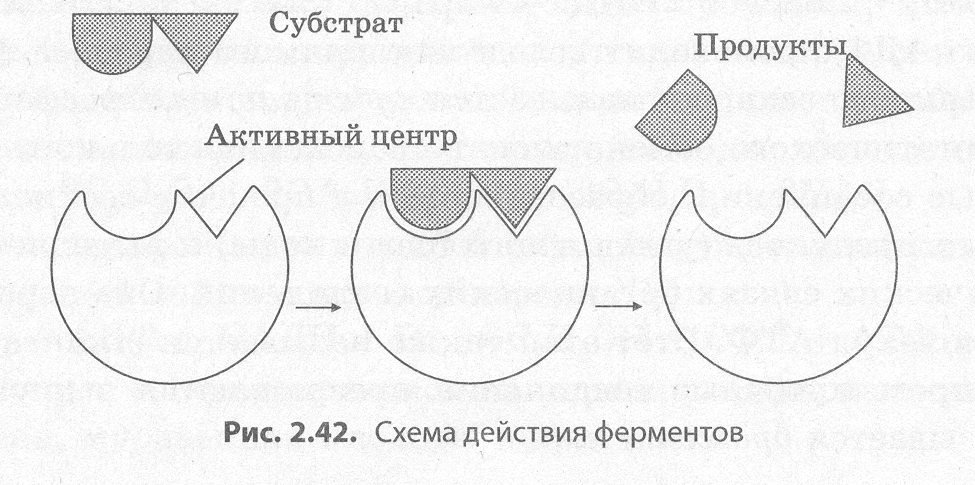
Протекание химических реакций в живых организмах обеспечивается благодаря биологиче�ским катализаторам белковой природы — ферментам, или энзимам. Как и другие катализаторы, ферменты ускоряют протекание химических реакций в клетке в десятки и сотни тысяч раз, а ино�гда и вообще делают их возможными, но не изменяют при этом ни природы, ни свойств конечно�го продукта (продуктов) реакции и не изменяются сами. Ферменты могут быть как простыми, так и сложными белками, в состав которых, кроме белковой части, входит и небелковая — кофактор (кофермент). Примерами ферментов являются амилаза слюны, расщепляющая полисахариды при длительном пережевывании, и пепсин, обеспечивающий переваривание белков в желудке.
Ферменты отличаются от катализаторов небелковой природы высокой специфичностью дей�ствия, значительным увеличением с их помощью скорости реакции, а также возможностью ре�гуляции действия за счет изменения условий протекания реакции либо взаимодействия с ними различных веществ. К тому же и условия, в которых протекает ферментный катализ, существен�но отличаются от тех, при которых идет неферментный: оптимальной для функционирования ферментов в организме человека является температура 37°С, давление должно быть близким к ат�мосферному, а рН среды может существенно колебаться. Так, для амилазы необходима щелочная среда, а для пепсина — кислая.
Механизм действия ферментов заключается в снижении энергии активации веществ (субстра�тов), вступающих в реакцию, за счет образования промежуточных фермент-субстратных ком�плексов (рис. 2.42).
Энергетический и пластический обмен, их взаимосвязь
Метаболизм складывается из двух одновременно протекающих в клетке процессов: пластиче�ского и энергетического обменов.
Пластический обмен (анаболизм, ассимиляция) представляет собой совокупность реакций синтеза, которые идут с затратой энергии АТФ. В процессе пластического обмена синтезируются органические вещества, необходимые клетке. Примером реакций пластического обмена являются фотосинтез, биосинтез белка и репликация (самоудвоение) ДНК.
Энергетический обмен (катаболизм, диссимиляция) — это совокупность реакций расщепле�ния сложных веществ до более простых. В результате энергетического обмена выделяется энер�гия, запасаемая в виде АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.
Пластический и энергетический обмены неразрывно связаны, поскольку в процессе пластиче�ского обмена синтезируются органические вещества и для этого необходима энергия АТФ, а в про�цессе энергетического обмена органические вещества расщепляются и высвобождается энергия, которая затем будет израсходована на процессы синтеза.
Энергию организмы получают в процессе питания, а высвобождают ее и переводят в доступную форму в основном в процессе дыхания. По способу питания все организмы делятся на автотрофов и гетеротрофов. Автотрофы способны самостоятельно синтезировать органические вещества из неорганических, а гетеротрофы используют исключительно готовые органические вещества.
Стадии энергетического обмена
Несмотря на всю сложность реакций энергетического обмена, его условно подразделяют на три этапа: подготовительный, анаэробный (бескислородный) и аэробный (кислородный).
На подготовительном этапе молекулы полисахаридов, липидов, белков, нуклеиновых кислот распадаются на более простые, например, глюкозу, глицерин и жирные кислоты, аминокислоты, нуклеотиды и др. Этот этап может протекать непосредственно в клетках либо в кишечнике, от�куда расщепленные вещества доставляются с током крови.
Анаэробный этап энергетического обмена сопровождается дальнейшим расщеплением моно�меров органических соединений до еще более простых промежуточных продуктов, например, пи- ровиноградной кислоты, или пирувата. Он не требует присутствия кислорода, и для многих ор�ганизмов, обитающих в иле болот или в кишечнике человека, является единственным способом получения энергии. Анаэробный этап энергетического обмена протекает в цитоплазме.
Бескислородному расщеплению могут подвергаться различные вещества, однако довольно ча�сто субстратом реакций оказывается глюкоза. Процесс ее бескислородного расщепления называет�ся гликолизом. При гликолизе молекула глюкозы теряет четыре атома водорода, т. е. окисляется, при этом образуются две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы восстановленного переносчика водорода НАДН + Н+:
С6Н1206 + 2Н3Р04 + 2АДФ + 2НАД → 2С3Н403 + 2АТФ + 2НАДН + Н+ + 2Н20.
Образование АТФ из АДФ происходит вследствие прямого переноса фосфат-аниона с предва�рительно фосфорилированного сахара и называется субстратным фосфорилированием.
Аэробный этап энергетического обмена может происходить только в присутствии кислорода, при этом промежуточные соединения, образовавшиеся в процессе бескислородного расщепления, окисляются до конечных продуктов (углекислого газа и воды) и выделяется большая часть энер�гии, запасенной в химических связях органических соединений. Она переходит в энергию макро- эргических связей 36 молекул АТФ. Этот этап также называется тканевым дыханием. В случае отсутствия кислорода промежуточные соединения превращаются в другие органические веще�ства, и этот процесс называется брожением.
Дыхание
Механизм клеточного дыхания схематически изображен на рис. 2.43.
Аэробное дыхание происходит в митохондриях, при этом пировиноградная кислота сначала утрачивает один атом углерода, что сопровождается синтезом одного восстановительного эквива�лента НАДН + Н+ и молекулы ацетилкофермента А (ацетил-КоА):
С3Н403 + НАД + Н
КоА →СН3СО
КоА + НАДН + Н+ + С02.
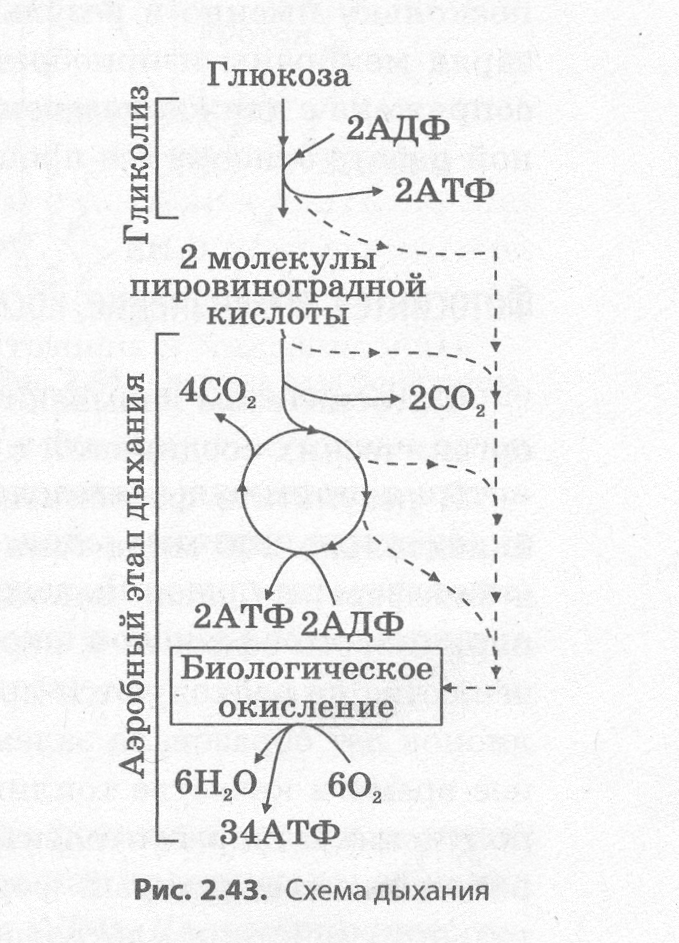
Ацетил-КоА в матриксе митохондрий вовлекается в цепь хими�ческих реакций, совокупность которых называется циклом Кребса (циклом трикарбоновых кислот, циклом лимонной кислоты). В хо�де этих превращений образуется две молекулы АТФ, ацетил-КоА полностью окисляется до углекислого газа, а его ионы водорода и электроны присоединяются к переносчикам водорода НАДН + Н+ и ФАДН2. Переносчики транспортируют протоны водорода и элек�троны к внутренним мембранам митохондрий, образующим кристы. При помощи белков-переносчиков протоны водорода нагнетаются в межмембранное пространство, а электроны передаются по так на�зываемой дыхательной цепи ферментов, расположенной на внутрен�ней мембране митохондрий, и сбрасываются на атомы кислорода:
02 +2е- →02-.
Следует отметить, что некоторые белки дыхательной цепи содер�жат железо и серу.
Из межмембранного пространства протоны водорода транспор�тируются обратно в матрикс митохондрий с помощью специальных ферментов — АТФ-синтаз, а выделяющаяся при этом энергия рас�ходуется на синтез 34 молекул АТФ из каждой молекулы глюкозы. Этот процесс называется окислительным фосфорилированием. В матриксе митохондрий протоны водорода реагируют с ра�дикалами кислорода с образованием воды:
4Н+ + О2-→2Н20.
Совокупность реакций кислородного дыхания может быть выражена следующим образом: 2С3Н403 + 602 + 36Н3Р04 + 36АДФ → 6C02 + 38Н20 + 36АТФ.
Суммарное уравнение дыхания выглядит таким образом:
С6Н1206 + 602 + 38Н3Р04 + 38АДФ→ 6С02 + 40Н20 + 38АТФ.
Брожение
В отсутствие кислорода или при его недостатке происходит брожение. Брожение является эволюционно более ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку в результате брожения образуются органические вещества, все еще богатые энергией. Различают несколько основных видов брожения: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода в ходе брожения пирови�ноградная кислота восстанавливается до молочной кислоты, при этом образовавшиеся ранее вос�становительные эквиваленты расходуются, и остаются всего две молекулы АТФ:
2С3Н403 + 2НАДН + Н+ → 2С3Н603 + 2НАД.
При брожении с помощью дрожжевых грибов пировиноградная кислота в присутствии кисло�рода превращается в этиловый спирт и оксид углерода (IV):
С3Н403 + Н3Р04 + АДФ + НАДН + Н+ →С2Н5ОН + С02 + АТФ + Н20 + НАД+ .
При брожении с помощью микроорганизмов из пировиноградной кислоты могут образоваться также уксусная, масляная, муравьиная кислоты и др.
АТФ, полученная в результате энергетического обмена, расходуется в клетке на различные виды работы: химическую, осмотическую, электрическую, механическую и регуляторную. Хими�ческая работа заключается в биосинтезе белков, липидов, углеводов, нуклеиновых кислот и дру�гих жизненно важных соединений. К осмотической работе относят процессы поглощения клеткой и выведения из нее веществ, которые во внеклеточном пространстве находятся в концентраци�ях, больших, чем в самой клетке. Электрическая работа тесно взаимосвязана с осмотической, поскольку именно в результате перемещения заряженных частиц через мембраны формируется заряд мембраны и приобретаются свойства возбудимости и проводимости. Механическая работа сопряжена с движением веществ и структур внутри клетки, а также клетки в целом. К регулятор- ной работе относят все процессы, направленные на координацию процессов в клетке.
Фотосинтез, его значение, космическая роль
Фотосинтезом называют процесс преобразования энергии света в энергию химических связей органических соединений с участием хлорофилла.
В результате фотосинтеза образуется около 150 млрд тонн органического вещества и при�близительно 200 млрд тонн кислорода ежегодно. Этот процесс обеспечивает круговорот углерода в биосфере, не давая накапливаться углекислому газу и препятствуя тем самым возникновению парникового эффекта и перегреву Земли. Образующиеся в результате фотосинтеза органические вещества не расходуются другими организмами полностью, значительная их часть в течение мил�лионов лет образовала залежи полезных ископаемых (каменного и бурого угля, нефти). В послед�нее время в качестве топлива начали использовать также рапсовое масло («биодизель») и спирт, полученный из растительных остатков. Из кислорода под действием электрических разрядов об�разуется озон, который формирует озоновый экран, защищающий все живое на Земле от губи�тельного действия ультрафиолетовых лучей.
Наш соотечественник, выдающийся физиолог растений К. А. Тимирязев (1843-1920) назвал роль фотосинтеза «космической», поскольку он связывает Землю с Солнцем (космосом), обеспечи�вая приток энергии на планету.
Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь
В 1905 году английский физиолог растений Ф. Блэкмен обнаружил, что скорость фотосинтеза не может увеличиваться беспредельно, какой-то фактор ограничивает ее. На основании этого он выдвинул предположение о наличии двух фаз фотосинтеза: световой и темновой. При низкой ин�тенсивности освещения скорость световых реакций возрастает пропорционально нарастанию силы света, и, кроме того, данные реакции не зависят от температуры, поскольку для их протекания не нужны ферменты. Световые реакции протекают на мембранах тилакоид.
Скорость темновых реакций, напротив, возрастает с повышением температуры, однако по достижении температурного порога в 30°С этот рост прекращается, что свидетельствует о фер�ментативном характере указанных превращений, происходящих в строме. Следует отметить, что свет также оказывает на темновые реакции определенное влияние, несмотря на то, что они на�зываются темновыми.
Световая фаза фотосинтеза (рис. 2.44) протекает на мембранах тилакоидов, несущих несколько типов белковых комплексов, основными из которых являются фотосистемы I и II, а также АТФ- синтаза. В состав фотосистем входят пигментные комплексы, в которых, кроме хлорофилла, при�сутствуют и каротиноиды. Каротиноиды улавливают свет в тех областях спектра, в которых этого не делает хлорофилл, а также защищают хлорофилл от разрушения светом высокой интенсивности.
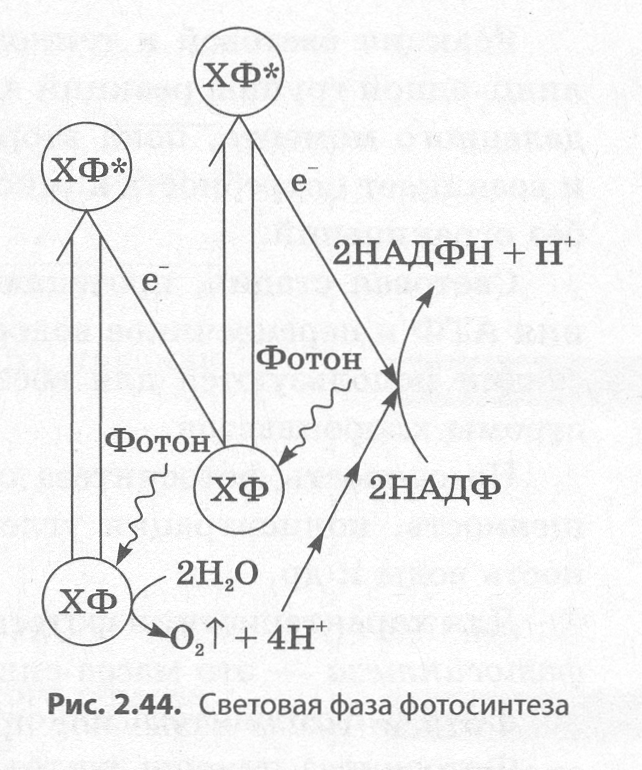
Кроме пигментных комплексов, фотосистемы включают и ряд белков-акцепторов электронов, которые последовательно передают друг другу электроны от молекул хлорифилла. Последователь�ность этих белков называется электронтранспортной цепью хлоропластов.
С фотосистемой II также ассоциирован специальный ком�плекс белков, который обеспечивает выделение кислорода в процессе фотосинтеза. Этот кислородвыделяющий комплекс содержит ионы марганца и хлора.
В световой фазе кванты света, или фотоны, попадающие на молекулы хлорофилла, расположенные на мембранах тилакоидов, переводят их в возбужденное состояние, характеризующе�еся более высокой энергией электронов. При этом возбужден�ные электроны от хлорофилла фотосистемы I передаются через цепь посредников на переносчик водорода НАДФ, присоединя�ющий при этом протоны водорода, всегда имеющиеся в водном растворе:
НАДФ + 2е-+ 2Н+ → НАДФН + Н+.
Восстановленный НАДФН + Н+ будет впоследствии исполь�зован в темновой стадии. Электроны от хлорофилла фотосисте�мы II также передаются по электронтранспортной цепи, однако они заполняют «электронные дырки» хлорофилла фотосистемы I. Недостаток электронов в хло�рофилле фотосистемы II заполняется за счет отнимания у молекул воды, которое происходит с участием уже упоминавшегося выше кислородвыделяющего комплекса. В результате разложе�ния молекул воды, которое называется фотолизом, образуются протоны водорода и выделяется молекулярный кислород, являющийся побочным продуктом фотосинтеза:
Н20 →2Н+ +2е- +1/2О2
Протоны водорода, накопившиеся в полости тилакоида в результате фотолиза воды и нагнета�ния при переносе электронов по электронтранспортной цепи, вытекают из тилакоида через канал в мембранном белке — АТФ-синтазе, при этом из АДФ синтезируется АТФ. Данный процесс на�зывается фотофосфорилированием. Он не требует участия кислорода, однако очень эффективен, так как дает в 30 раз больше АТФ, чем митохондрии в процессе окисления. Образовавшаяся в све�товых реакциях АТФ впоследствии будет использована в темновых реакциях.
Суммарное уравнение реакций световой фазы фотосинтеза можно записать следующим обра�зом:
2Н20 + 2НАДФ + 3АДФ + ЗН3Р04 → 2НАДФН + Н+ + 3АТФ.
В ходе темновых реакций фотосинтеза (рис. 2.45) происходит связывание молекул С02 в виде углеводов, на которое расходуются молекулы АТФ и НАДФН + Н+, синтезированные в световых реакциях:
6С02 + 12 НАДФН + Н+ + 18АТФ→ С6Н1206 + 6Н20 + 12 НАДФ + 18АДФ + 18Н3Р04.
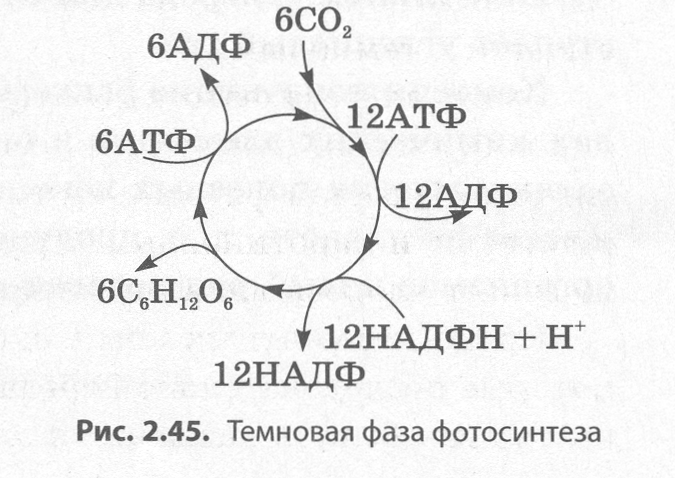
Процесс связывания углекислого газа является сложной цепью превращений, названной ци�клом Кальвина в честь его первооткрывателя. Темновые реакции протекают в строме хлоропластов. Для их протекания необходим постоянный приток углекислого газа извне через устьица, а затем и по системе межклетников.
Первыми в процессе фиксации углекислого газа образуются трехуглеродные сахара, являю�щиеся первичными продуктами фотосинтеза, тогда как образующуюся позже глюкозу, которая расходуется на синтез крахмала и другие процессы жизнедея�тельности, называют конечным продуктом фотосинтеза.
Таким образом, в процессе фотосинтеза энергия солнечного света преобразуется в энергию химических связей сложных ор�ганических соединений не без участия хлорофилла. Суммарное уравнение фотосинтеза можно записать следующим образом:
6С02 + 12Н20 → С6Н1206 + 602 + 6Н20, или
6С02 + 6Н20 →С6Н1206 + 602.
Реакции световой и темновой фаз фотосинтеза взаимосвязаны, так как увеличение скорости лишь одной группы реакций влияет на интенсивность всего процесса фотосинтеза только до опре�деленного момента, пока вторая группа реакций не выступит в роли лимитирующего фактора, и возникает потребность в ускорении реакций второй группы для того, чтобы первые происходили без ограничений.
Световая стадия, протекающая в тилакоидах, обеспечивает запасание энергии для образова�ния АТФ и переносчиков водорода. На второй стадии, темновой, энергетические продукты первой стадии используются для восстановления углекислого газа, и происходит это в компартментах стромы хлоропластов.
На скорость фотосинтеза оказывают влияние различные факторы окружающей среды: осве�щенность, концентрация углекислого газа в атмосфере, температура воздуха и почвы, доступ�ность воды и др.
Для характеристики фотосинтеза используется понятие его продуктивности.
Продуктивность фотосинтеза — это масса синтезируемой за 1 час глюкозы на 1 дм2 листовой поверхности. Этот показатель фотосинтеза максимален при оптимальных условиях.
Фотосинтез присущ не только зеленым растениям, но и многим бактериям, в том числе ци- анобактерям, зеленым и пурпурным бактериям, однако у последних он может иметь некоторые отличия, в частности, при фотосинтезе бактерии могут не выделять кислорода (это не касается цианобактерий).
Хемосинтез. Роль хемосинтезирующих бактерий на Земле
Хемосинтез — это процесс синтеза органических соединений за счет химической энергии не�органических соединений.
Данный процесс был открыт выдающимся русским ученым С.Н. Виноградским в 1887 го�ду. К группе хемосинтетиков (хемотрофов) относятся в основном бактерии: нитрифицирующие, серобактерии, железобактерии и др. Они используют энергию окисления соединений азота, серы, ионов железа соотвественно. При этом донором электронов выступает не вода, а другие неоргани�ческие вещества.
Так, нитрифицирующие бактерии окисляют образованный из атмосферного азота азотфиксирующими бактериями аммиак до нитритов и нитратов:
2NH3 +302 → 2HNO2 + 2Н20 + 663 кДж,
2HN02 + 02 →2HN03 + 192 кДж.
Серобактерии окисляют сероводород до серы, а в некоторых случаях и до серной кислоты:
H2S + 02 → 2Н20 + 2S + 272 кДж,
2S + 302 + Н20 → H2S04 + 483 кДж.
Железобактерии окисляют соли железа:
4FeC03 + 02 + 6Н20 →4Fe(OH)3 + 4С02 + 324 кДж.
Водородные бактерии способны окислять молекулярный водород:
2Н2 + 02 → 2Н20 + 235 кДж.
Источником углерода для синтеза органических соединений у всех автотрофных бактерий вы�ступает углекислый газ.
Хемосинтезирующие бактерии наиболее значительную роль играют в биогеохимических цик�лах химических элементов в биосфере, так как в процессе их жизнедеятельности образовались залежи многих полезных ископаемых. Кроме того, они являются источниками органического ве�щества на планете, т. е. продуцентами, а также делают доступным и для растений, и для других организмов целый ряд неорганических веществ.
2.6. Биосинтез белка и нуклеиновых кислот. Матричный характер реакций биосинтеза. Генетическая информация в клетке. Гены, генетический код и его свойства.
Генетическая информация в клетке
Воспроизведение себе подобных является одним из фундаментальных свойств живого. Благо�даря этому явлению существует сходство не только между организмами, но и между отдельными клетками, а также их органоидами (митохондриями и пластидами). Материальной основой этого сходства является передача зашифрованной в последовательности нуклеотидов ДНК генетической информации, которая осуществляется благодаря процессам репликации (самоудвоения) ДНК. Реализуются все признаки и свойства клеток и организмов благодаря белкам, структуру которых в первую очередь и определяют последовательности нуклеотидов ДНК. Поэтому первостепенное значение в процессах метаболизма играет именно биосинтез нуклеиновых кислот и белка. Струк�турной единицей наследственной информации является ген.
Гены, генетический код и его свойства
Наследственная информация в клетке не является монолитной, она разбита на отдельные «слова» — гены.
Ген — это элементарная единица генетической информации.
Работы по программе «Геном человека», которые проводились одновременно в нескольких странах и были завершены в начале нынешнего века, дали нам понимание того, что у человека всего около 25-30 тыс. генов, но информация с большей части нашей ДНК не считывается ни�когда, так как в ней содержится огромное количество бессмысленных участков, повторов и генов, кодирующих признаки, утратившие значение для человека (хвост, оволосение тела и др.). Кроме того, был расшифрован ряд генов, отвечающих за развитие наследственных заболеваний, а также генов-мишеней лекарственных препаратов. Однако практическое применение результатов, полу�ченных в ходе реализации данной программы, откладывается до тех пор, пока не будут расшиф�рованы геномы большего количества людей и станет понятно, чем же все-таки они различаются.
Гены, кодирующие первичную структуру белка, рибосомальной или транспортной РНК на�зываются структурными, а гены, обеспечивающие активацию или подавление считывания ин�формации со структурных генов, — регуляторными. Однако даже структурные гены содержат регуляторные участки.
Наследственная информация организмов зашифрована в ДНК в виде определенных сочетаний нуклеотидов и их последовательности — генетического кода. Его свойствами являются: триплет- ность, специфичность, универсальность, избыточность и неперекрываемость. Кроме того, в гене�тическом коде отсутствуют знаки препинания.
Каждая аминокислота закодирована в ДНК тремя нуклеотидами — триплетом, например, метионин закодирован триплетом ТАЦ, то есть код триплетен (табл. 2.4). С другой стороны, каж�дый триплет кодирует только одну аминокислоту, в чем заключается его специфичность или одно�значность. Генетический код универсален для всех живых организмов, то есть наследственная информация о белках человека может считываться бактериями и наоборот. Это свидетельствует о единстве происхождения органического мира. Однако 64 комбинациям нуклеотидов по три со�ответствует только 20 аминокислот, вследствие чего одну аминокислоту может кодировать 2-6 триплетов, то есть генетический код избыточен, или вырожден. Три триплета не имеют соответ�ствующих аминокислот, их называют стоп-кодонами, так как они обозначают окончание синтеза полипептидной цепи.
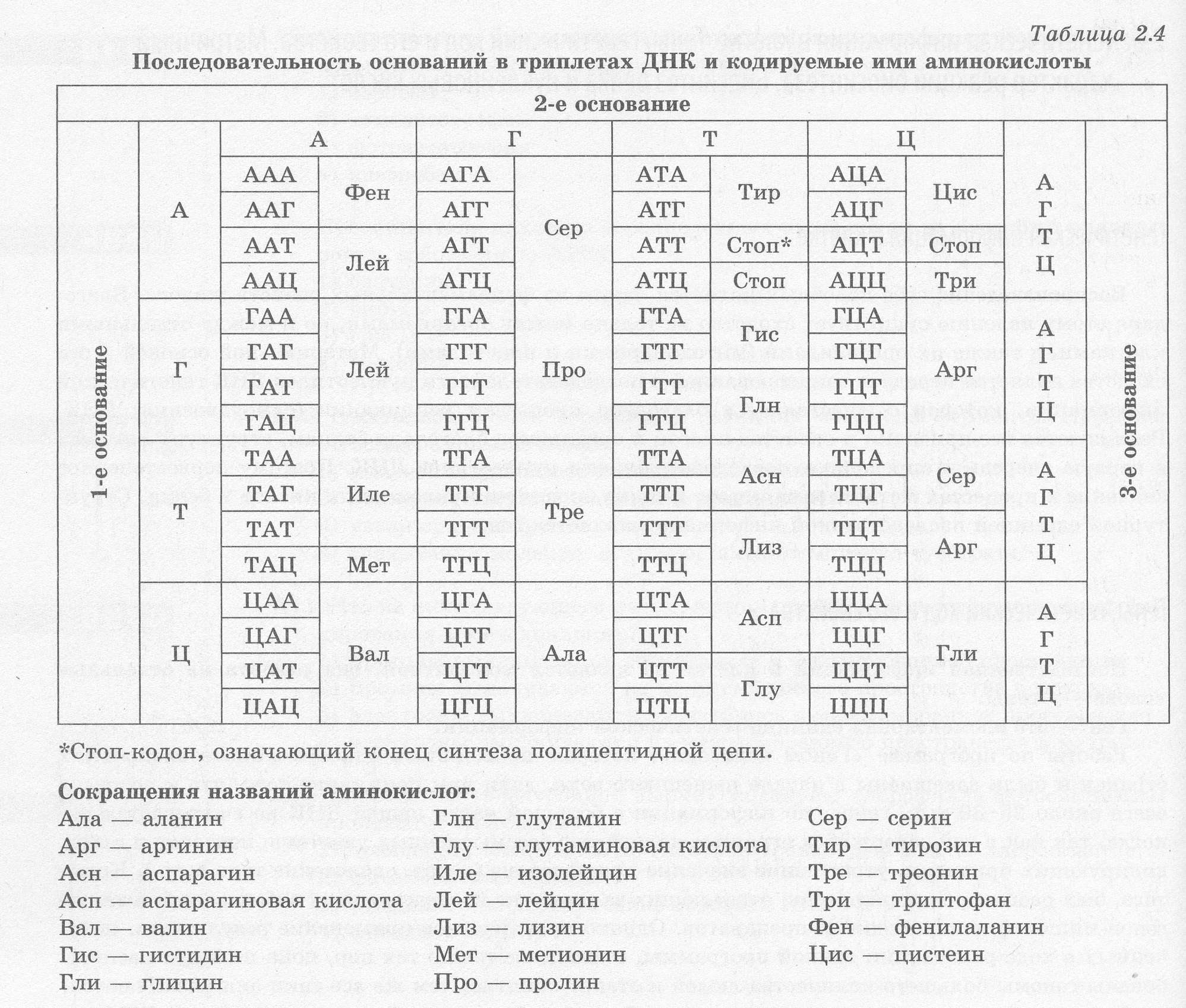
Если начать считывание генетической информации не с первого нуклеотида в триплете, а со второго, то произойдет не только сдвижка рамки считывания — синтезированный таким обра�зом белок будет совсем иным не только по последовательности нуклеотидов, но и по структуре и свойствам. Между триплетами отсутствуют какие бы то ни было знаки препинания, поэтому нет никаких препятствий для сдвижки рамки считывания, что открывает простор для возникновения и сохранения мутаций.
Матричный характер реакций биосинтеза
Клетки бактерий способны удваиваться каждые 20-30 минут, а клетки эукариот — каждые сутки и даже чаще, что требует высокой скорости и точности репликации ДНК. Кроме того, каж�дая клетка содержит сотни и тысячи копий многих белков, особенно ферментов, следовательно, для их воспроизведения неприемлем «штучный» способ их производства. Более прогрессивным способом является штамповка, которая позволяет получить многочисленные точные копии про�дукта и к тому же снизить его себестоимость. Для штамповки необходима матрица, с которой осуществляется оттиск.
В клетках принцип матричного синтеза заключается в том, что новые молекулы белков и ну�клеиновых кислот синтезируются в соответствии с программой, заложенной в структуре ранее существовавших молекул тех же нуклеиновых кислот (ДНК или РНК).
Биосинтез белка и нуклеиновых кислот
Репликация ДНК. ДНК представляет собой двухцепочечный биополимер, мономерами которо�го являются нуклеотиды. Если бы биосинтез ДНК происходил по принципу ксерокопирования, то неизбежно возникали бы многочисленные искажения и погрешности в наследственной информа�ции, которые в конечном итоге привели к гибели новых организмов. Поэтому процесс удвоения ДНК происходит иным, полуконсервативным способом: молекула ДНК расплетается, и на каж�дой из цепей синтезируется новая цепь по принципу комплементарности. Процесс самовоспроиз�ведения молекулы ДНК, обеспечивающий точное копирование наследственной информации и пе�редачу ее из поколения в поколение, называется репликацией (от лат. репликацио — повторение). В результате репликации образуются две абсолютно точные копии материнской молекулы ДНК, каждая из кото�рых несет по одной копии материнской (рис. 2.46).
Процесс репликации на самом деле крайне сложен, так как в нем участвует целый ряд белков. Одни из них раскручивают двойную спи�раль ДНК, другие разрывают водородные связи между нуклеотидами комплементарных цепей, третьи (например, фермент ДНК-полимераза) подбирают по принципу комплементарности новые нуклеотиды и т. д. Образовавшиеся в результате репликации две молекулы ДНК в процес�се деления расходятся по двум вновь образующимся дочерним клеткам.
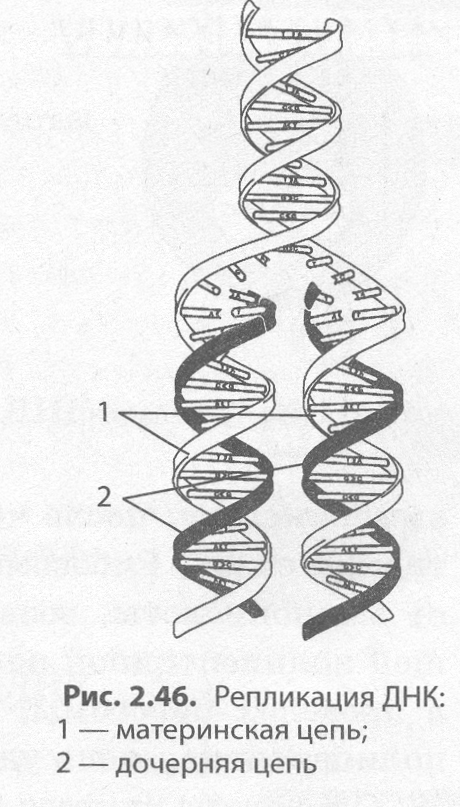
Ошибки в процессе репликации возникают крайне редко, однако если они и происходят, то очень быстро устраняются как ДНК- полимеразами, так и специальными ферментами репарации, поскольку любая ошибка в последовательности нуклеотидов может привести к не�обратимому изменению структуры и функций белка и, в конечном ито�ге, неблагоприятно сказаться на жизнеспособности новой клетки или даже особи.
Биосинтез белка. Как образно выразился выдающийся философ XIX века Ф. Энгельс: «Жизнь есть форма существования белковых тел». Структура и свойства белковых молекул определяются их первичной структурой, т. е. последовательностью аминокислот, зашифрованной в ДНК. От точности воспроизведения этой информации зависит не только существование самого полипепти�да, но и функционирование клетки в целом, поэтому процесс синтеза белка имеет огромное зна�чение. Он, по-видимому, является самым сложным процессом синтеза в клетке, поскольку здесь участвует до трехсот различных ферментов и других макромолекул. Кроме того, он протекает с высокой скоростью, что требует еще боль�шей точности.
В биосинтезе белка выделяют два основных этапа: транскрипцию и трансляцию.
Транскрипция (от лат. транскрипций — переписывание) — это био�синтез молекул иРНК на соответствующих участках ДНК (рис. 2.47).
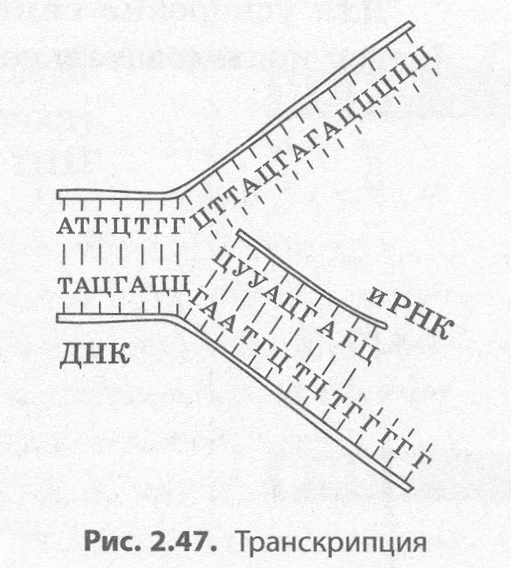
Поскольку молекула ДНК содержит две антипараллельных цепи, то считывание информации с обеих цепей привело бы к образованию совершенно различных иРНК, поэтому их биосинтез возможен только на одной из цепей, которую называют кодирующей, в отличие от вто�рой, некодирующей. Обеспечивает процесс переписывания специаль�ный фермент РНК-полимераза, который подбирает нуклеотиды РНКпо принципу комплементарности. Этот процесс может протекать как в ядре, так и в органоидах, имеюших собственную ДНК, — митохондриях и пластидах.
Синтезированные в процессе транскрипции молекулы иРНК покидают ядро через ядерные поры и проходят в цитоплазме сложный процесс подготовки к трансляции (митохондриальные и пластидные иРНК могут оставаться внутри органоидов, где и происходит второй этап биосин�теза белка). В процессе созревания иРНК к ней присоединяются первые три нуклеотида (АУГ) и хвост из адениловых нуклеотидов, длина которого определяет, сколько копий белка может син�тезироваться на данной молекуле.
После транскрипции происходит процесс активации аминокислот, в ходе которого аминокис�лота присоединяется к соответствующей свободной тРНК. Этот процесс катализируется специаль�ным ферментом, на него затрачивается АТФ.
Трансляция (от лат. трансляций — передача) — это биосинтез полипептидной цепи на моле�куле иРНК, при котором происходит перевод генетической информации в последовательность аминокислот полипептидной цепи (рис. 2.48).
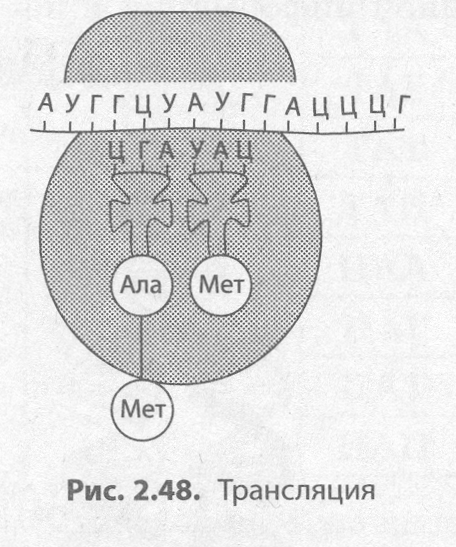
Второй этап синтеза белка чаще всего происходит в цитоплазме, на�пример на шероховатой ЭПС. Для его протекания необходимы наличие рибосом, активация тРНК, в ходе которой они присоединяют соответ�ствующие аминокислоты, присутствие ионов Mg2+, а также оптималь�ные условия среды (температура, рН, давление и т. д.).
Для начала транскрипции (инициации) к готовой к синтезу моле�куле иРНК присоединяется малая субъединица рибосомы, а затем по принципу комплементарности к первому кодону подбирается тРНК, несущая аминокислоту метионин. Лишь после этого присоединяет�ся большая субъединица рибосомы. В пределах собранной рибосомы оказываются два кодона иРНК, первый из которых уже занят. К со�седнему с ним кодону присоединяется вторая тРНК, также несущая аминокислоту, после чего между остатками аминокислот с помощью ферментов образуется пеп�тидная связь. Рибосома передвигается на один кодон иРНК; первая из тРНК, освободившаяся от аминокислоты, возвращается в цитоплазму за следующей аминокислотой, а фрагмент буду�щей полипептидной цепи как бы повисает на оставшейся тРНК. К новому кодону, оказавшемуся в пределах рибосомы, присоединяется следующая тРНК, процесс повторяется и шаг за шагом полипептидная цепь удлиняется, т. е. происходит ее элонгация.
Окончание синтеза белка (терминация) происходит, как только в молекуле иРНК встретится специфическая последовательность нуклеотидов, которая не кодирует аминокислоту (стоп-кодон). После этого рибосома, иРНК и полипептидная цепь разделяются, а вновь синтезированный белок приобретает соответствующую структуру и транспортируется в ту часть клетки, где он будет вы�полнять свои функции.
Трансляция является весьма энергоемким процессом, поскольку на присоединение одной ами�нокислоты к тРНК расходуется энергия одной молекулы АТФ, еще несколько используются для продвижения рибосомы по молекуле иРНК.
Для ускорения синтеза определенных белковых молекул к молекуле иРНК могут присоеди�няться последовательно несколько рибосом, которые образуют единую структуру — полисому.
2.7. Клетка – генетическая единица живого. Хромосомы, их строение (форма и размеры) и функции. Число хромосом и их видовое постоянство. Особенности соматических и половых клеток. Жизненный цикл клетки: интерфаза и митоз. Митоз – деление соматических клеток. Мейоз. Фазы митоза и мейоза. Развитие половых клеток у растений и животных. Сходство и отличие митоза и мейоза, их значение. Деление клетки – основа роста, развития и размножения организмов. Роль мейоза в обеспечении постоянства числа хромосом в поколениях.
Клетка — генетическая единица живого
Несмотря на то, что нуклеиновые кислоты являются носителем генетической информации, реа�лизация этой информации невозможна вне клетки, что легко доказывается на примере вирусов. Данные организмы, содержащие зачастую только ДНК или РНК, не могут самостоятельно воспро�изводиться, для этого они должны использовать наследственный аппарат клетки. Даже проник�нуть в клетку без помощи самой клетки они не могут, кроме как с использованием механизмов мембранного транспорта или благодаря повреждению клеток. Большинство вирусов нестабильно, они гибнут уже после нескольких часов пребывания на открытом воздухе. Следовательно, клетка является генетической единицей живого, обладающей минимальным набором компонентов для со�хранения, изменения и реализации наследственной информации, а также ее передачи потомкам.
Большая часть генетической информации эукариотической клетки сосредоточена в ядре. Осо�бенностью ее организации является то, что, в отличие от ДНК прокариотической клетки, молеку�лы ДНК эукариот не замкнуты и образуют сложные комплексы с белками — хромосомы.
Хромосомы, их строение (форма и размеры) и функции
Хромосома (от греч. хрома — цвет, окраска и сома — тело) — это структура клеточного ядра, которая содержит гены и несет определенную наследственную информацию о признаках и свой�ствах организма.
Иногда хромосомами называют и кольцевые молекулы ДНК прокариот. Хромосомы способны к самоудвоению, они обладают структурной и функциональной индивидуальностью и сохраняют ее в ряду поколений. Каждая клетка несет всю наследственную информацию организма, но в ней работает только небольшая часть.
Основой хромосомы является двухцепочечная молекула ДНК, упакованная с белками. У эу�кариот с ДНК взаимодействуют гистоновые и негистоновые белки, тогда как у прокариот гистоновые белки отсутствуют.
Лучше всего хромосомы видны под световым микроскопом в про�цессе деления клетки, когда они в результате уплотнения приобретают вид палочковидных телец, разделенных первичной перетяжкой — цен�тромерой — на плечи. На хромосоме может быть также и вторичная перетяжка, которая в некоторых случаях отделяет от основной части хромосомы так называемый спутник. Концевые участки хромосом на�зываются теломерами. Теломеры препятствуют слипанию концов хро�мосом и обеспечивают их прикрепление к оболочке ядра в неделящейся клетке. В начале деления хромосомы удвоены и состоят из двух дочер�них хромосом — хроматид, скрепленных в центромере (рис. 2.49).
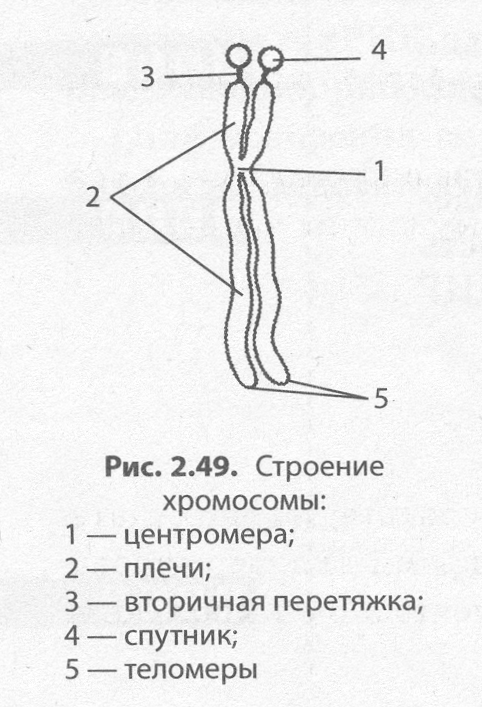
По форме различают равноплечие, неравноплечие и палочковидные хромосомы. Размеры хромосом существенно варьируют, однако средняя хромосома имеет размеры 5 х 1,4 мкм.
В некоторых случаях хромосомы в результате многочисленных удвоений ДНК содержат сотни и тысячи хроматид: такие гигантские хромосомы называются политенными. Они встречаются в слюнных железах личинок дрозофилы, а также в пищеварительных железах аскариды.
Число хромосом и их видовое постоянство. Соматические и половые клетки
Согласно клеточной теории клетка является единицей строения, жизнедеятельности и раз�вития организма. Таким образом, такие важнейшие функции живого, как рост, размножение и развитие организма обеспечиваются на клеточном уровне. Клетки многоклеточных организмов можно разделить на соматические и половые.
Соматические клетки — это все клетки тела, образующиеся в результате митотического деле�ния.
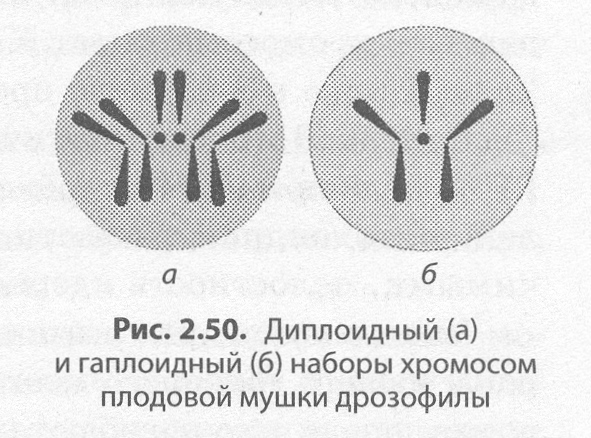
Изучение хромосом позволило установить, что для соматических клеток организма каждого биологического вида характерно постоянное число хромосом. Например, у человека их 46. Набор хромосом соматических клеток называют диплоидным (2п), или двойным (рис. 2.50, а).
Половые клетки, или гаметы, — это специализированные клетки, служащие для полового размножения.
В гаметах содержится всегда вдвое меньше хромосом, чем в соматических клетках (у человека — 23), поэтому набор хромо�сом половых клеток называется гаплоидным (п), или одинарным (рис. 2.50, б). Его образование связано с мейотическим делением клетки.
Количество ДНК соматических клеток обозначается как 2с, а половых — 1с. Генетическая формула соматических клеток за�писывается как 2п2с, а половых — 1n1c.
В ядрах некоторых соматических клеток количество хромосом может отличаться от их коли�чества в соматических клетках. Если это различие больше на один, два, три и т. д. гаплоидных набора, то такие клетки называют полиплоидными (три-, тетра-, пентаплоидными соответствен�но). В таких клетках процессы метаболизма протекают, как правило, очень интенсивно.
Количество хромосом само по себе не является видоспецифическим признаком, поскольку раз�личные организмы могут иметь равное количество хромосом, а родственные — разное. Например, у малярийного плазмодия и лошадиной аскариды по две хромосомы, а у человека и шимпанзе — 46 и 48 соответственно.
Хромосомы человека делятся на две группы: аутосомы и половые хромосомы (гетерохромосо- мы). Аутосом в соматических клетках человека насчитывается 22 пары, они одинаковы для муж�чин и женщин, а половых хромосом только одна пара, но именно она определяет пол особи. Су�ществует два вида половых хромосом — X и Y. Клетки тела женщины несут по две Х-хромосомы, а мужчин — X и Y.
Кариотип — это совокупность признаков хромосомного набора организма (число хромосом, их форма и величина).
Условная запись кариотипа включает общее количество хромосом, половые хромосомы и воз�можные отклонения в наборе хромосом. Например, кариотип нормального мужчины записывает�ся как 46, XY, а кариотип нормальной женщины — 46, XX.
Жизненный цикл клетки: интерфаза и митоз
Клетки не возникают каждый раз заново, они образуются только в результате деления мате�ринских клеток. После разделения дочерним клеткам требуется некоторое время для формирова�ния органоидов и приобретения соответствующей структуры, которая обеспечила бы выполнение определенной функции. Этот отрезок времени называется созреванием.
Промежуток времени от появления клетки в ре�зультате деления до ее разделения или гибели на�зывается жизненным циклом клетки.
У эукариотических клеток жизненный цикл де�лится на две основные стадии: интерфазу и митоз (рис. 2.51).
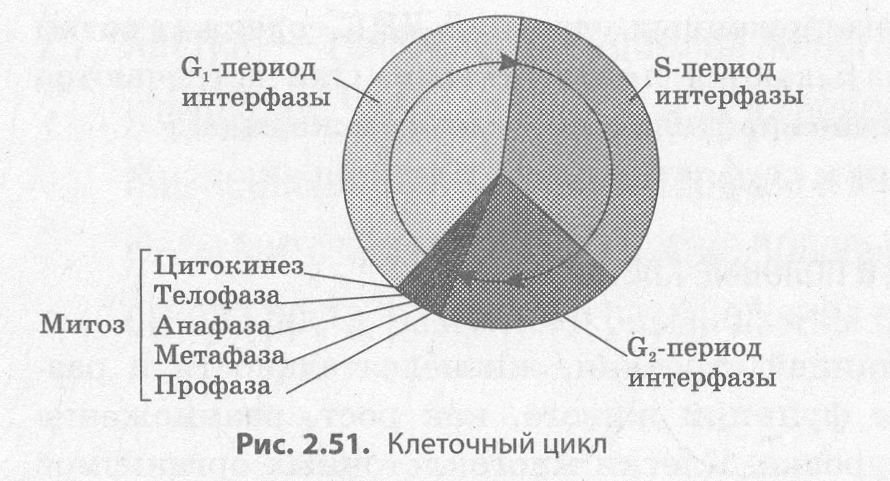
Интерфаза — это промежуток времени в жиз�ненном цикле, в который клетка не делится и нор�мально функционирует. Интерфаза делится на три периода: G1, S- и G 2-периоды.
G1 -период (пресинтетический, постмитотический) — это период роста и развития клетки, в ко�торый происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь образовавшейся клетки. К концу этого периода клетка может начать готовиться к удвоению ДНК.
ВS-nepuode (синтетическом) происходит сам процесс репликации ДНК. Единственным участ�ком хромосомы, который не подвергается репликации, является центромера, поэтому образо�вавшиеся молекулы ДНК не расходятся полностью, а остаются скрепленными в ней, и в начале деления хромосома имеет Х-образный вид. Генетическая формула клетки после удвоения ДНК — 2п4с. Также в S-периоде происходит удвоение центриолей клеточного центра.
G2-nepuoд (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центрио�лей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо разли�чимыми, целостность ядерной оболочки не нарушается, а органоиды не изменяются.
|
|
|
 Скачать 22.74 Mb.
Скачать 22.74 Mb.