Коллоквиум по физиологии. Колоквиум 2 ЦНС. Коллоквиум по разделу Общая и частная физиология цнс
 Скачать 1.11 Mb. Скачать 1.11 Mb.
|

|
Часть волокон, проводящих тактильную чувствительность, не образует перекреста и следует в головной мозг в заднем канатике вместе с тонким и клиновидным пучками. Передний и латеральный спиноталамические пути нередко объединяют в один спиноталамический пучок, в котором волокна, идущие от рецепторов, воспринимающих давление, проходят в переднем канатике ближе к средней линии. Латеральнее расположены волокна, проводящие чувство осязания, а затем проводящие чувство боли и температуру. К этой же группе относятся проводящие пути органов чувств. Передний и задний спиномозжечковые пути – пути Говерса и Флексига – афферентные пути мышечно - суставной чувствительности мозжечкового направления. Путь двухнейронный. Тела 1-х нейронов располагаются в соответствующих спинномозговых узлах. Их дендриты направляются на периферию в составе спинно-мозговых нервов и заканчиваются проприорецепторами в надкостнице, связках, суставной капсуле, сухожилиях, мышцах. Аксоны в составе задних корешков вступают в СМ и идут к промежуточной зоне. Здесь в nucleus intermediomedialis лежат 2-е нейроны. Аксоны клеток 2-го нейрона переходят через переднюю серую спайку в боковой канатик противоположной стороны, и поднимаются вверх до перешейка ромбовидного мозга. В этом месте волокна этого пути возвращаются на свою сторону и через верхнюю мозжечковую ножку вступают в кору червя мозжечка своей стороны. Проприоцептивные импульсы, поступившие в кору червя по переднему спинно-мозжечковому тракту, также передаются в красное ядро и через зубчатое ядро в кору большого мозга. Задний спинно-мозжечковый путь Флексига, двухнейронный. 1-ый нейрон – в спинальном ганглии. Дендриты оканчиваются в надкостнице. Аксоны оканчиваются на нейронах грудного ядра (2 нейрон). Его аксоны образуют задний спинно-мозжечковый путь. Путь проходит в боковом канатике, не перекрещивается до коры червя. Проприоцептивные путимозжечкового направления и связанные с корой больших полушарий дают возможность человеку (особенно спортсменам) тонко чувствовать свое тело и точно выполнять высокодифференцированные движения. Обработка различных физических упражнений сопровождается в первую очередь, тренировкой нервной системы, а затем уже двигательного аппарата. Тонкий пучок Голля и клиновидный пучок Бурдаха – афферентные пути мышечно – суставной чувствительности коркового направления от нижний конечностей и нижней половины тела и от верхний конечностей и верхней половины тела соответственно. Рецепторы этого вида чувствительности (проприорецепторы) располагаются в мышцах, сухожилиях и надкостнице. Рецепторами сухожилий являются тельца Гольджи (возбуждаются при растяжении). В фасциях находятся тельца Пачини (возбуждаются при давлении). Это сознательный, восходящий, трехнейронный путь. Тела первых нейронов находятся в клетках межпозвоночных узлов, их дендриты заканчиваются рецепторами, а аксоны входят в задние корешки и направляются в задние столбы. Волокна, идущие от ног, располагаются медиально, образуя пучок Голля (нежный), а идущие от рук — латерально, образуя пучок Бурдаха (клиновидный). В составе указанных пучков аксоны тел вторых нейронов доходят до ядер задних канатиков (Голля и Бурдаха), где начинаются вторые нейроны, их аксоны совершают перекрест в межоливном слое и заканчиваются в вентролатеральных ядрах таламусов, откуда начинаются третьи нейроны, проходят задний отдел задней ножки внутренних капсул и в составе лучистых венцов направляются в проекционную чувствительную зону — задние центральные извилины (поля 1, 2, 3), верхние теменные (поля 5—7), а также по ассоциативным волокнам в передние центральные извилины (поля 4, 6). 19. Физиология таламуса. Функциональная характеристика и особенности его ядерных групп. Таламус, или зрительный бугор, является одним из важнейших образований головного мозга, он тесно связан морфологически и функционально с корой больших полушарий. Не без основания считается, что в таламусе хранятся ключи к тайнам коры головного мозга. Деятельность таламуса тесно связана с анализом афферентных сигналов, организацией интегративных процессов, он участвует в механизмах регуляции функционального состояния и высшей нервной деятельности. Специфические, или релейные переключательные, ядра таламуса характеризуются следующими особенностями. Имеют локальную проекцию в строго определенные участки коры. Моносинаптически связаны аксосоматическими синапсами с нейронами третьего-четвертого слоев коры. В них переключается афферентная импульсация от периферических рецепторов или от первичных воспринимающих ядер нижележащих стволовых структур, а также от внесенсорных источников. Основную массу клеток составляют «релейные» «Релейные* клетки окружены вставочными интернейронами, многие из которых имеют тормозную функцию. К специфическим ядрам таламуса относят переднее дорсальное, переднее вентральное, переднее медиальное, вентральное постлатеральное, вентральное постмедиальное, вентральное латеральное, а также латеральное и медиальное коленчатые тела. В функциональном отношении специфические ядра представляют собой важнейшую часть основных сенсорных и моторных систем, разрушение релейных ядер приводит к полной и необратимой потере соответствующей чувствительности или нарушениям движений. Неспецифические ядра таламуса не относятся к какой-либо одной специфической проекционной или ассоциативной системе: они морфологически и функционально связаны со многими системами и участвуют вместе с ретикулярной формацией мозгового ствола в осуществлении неспецифических функций. Неспецифические ядра таламуса характеризуются следующими особенностями. Проецируются на кору больших полушарий более диффузно, чем специфические ядра. Связь с корой в основном полисинаптическая; волокна направляются к нейронам всех слоев коры и образуют аксодендритические синапсы на пирамидных нейронах. Афферентация к ним поступает, главным образом, из ретикулярной формации мозгового ствола, а также гипоталамуса, лимбической системы, базальных ядер, специфических ядер таламуса, имеются и отдельные коллатерали от сенсорных путей. Нейронные сети имеют преимущественно «ретикулярное» строение, т.е. состоят, в основном, из густой сети нейронов с длинными, слабо ветвящимися дендритами. К неспецифическим ядрам таламуса относят: срединный центр, парацентральное ядро, центральное медиальное, центральное латеральное, субмедиалъ-ное, вентральное переднее, парафасцикулярный комплекс, ретикулярное, перивентикулярное и центральную серую массу. Деятельность неспецифических ядер таламуса приводит к генерации «веретенообразной* электрической активности в таламусе и коре. Ассоциативные ядра таламуса наиболее поздно дифференцируются и интенсивно развиваются в ходе эволюции зрительного бугра. Они характеризуются следующими признаками. Эфферентные волокна направляются к ассоциативным областям коры, а также частично и к специфическим проекционным областям. Связь с ассоциативными областями коры моносинаптическая; волокна идут к I и II слоям коры, отдавая по пути коллатерали в IV и V слои и образуя аксоматические контакты с пирамидными нейронами. К ассоциативным ядрам относятся: медиодорсальное, латеральное дорсальное и подушка.. Важной особенностью нейронов ассоциативных ядер является способность многих из них реагировать на раздражения разных сенсорных входов, большая часть нейронов является двух-или полисенсорными. 20. Характеристика статических рефлексов. Динамика их организации. Статические рефлексы возникают при изменении положения тела и его частей в пространстве: 1.при изменении положения головы в пространстве — это так называемые лабиринтные рефлексы, возникающие в результате раздражения рецепторов вестибулярного аппарата; 2.при изменении положения головы по отношению к туловищу — шейные рефлексы с проприореоров мыщц шеи; 3.при нарушении нормальной позы тела —вырямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища, которые обеспечивают возвращение тела в вертикальное положение. Макулярные органы осуществляют статические рефлексы, которые поддерживают равновесие при разнообразных стоячих и наклонных положениях тела. Компенсаторное вращение глаз, особенно хорошо заметное у кошек, но возникающее и у людей, представляет собой статический рефлекс. Благодаря ему на сетчатке сохраняется изображение горизонтальных и вертикальных линий. Статические рефлексы обеспечивают перераспределение тонуса мышц, для сохранения позы и равновесия. 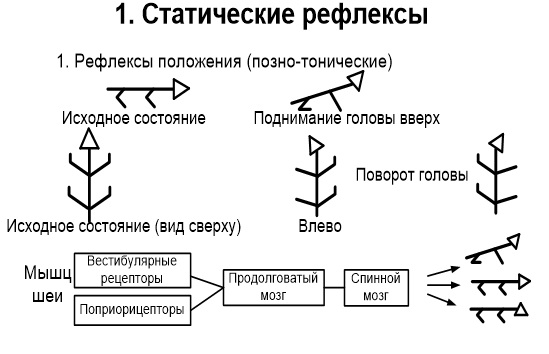 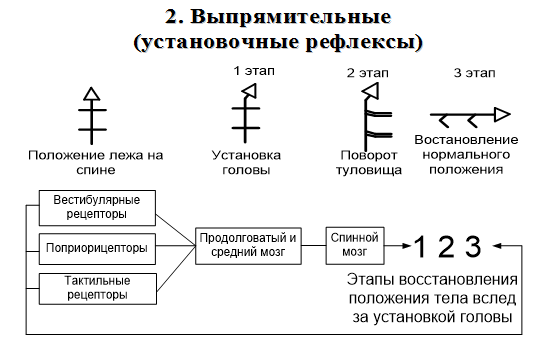 21. Характеристика статокинетических рефлексов. Динамика их организации. Статокинетического рефлексы направлены на сохранение позы и поддержания равновесия в случае ускорения. Различают горизонтальное (начало или окончание движения в этой плоскости), вертикальное (например, так называемые лифтни реакции) и угловое (при поворотах) ускорение. При этом раздражаются волосковые клетки полукружных каналов. Оттуда вестибулослуховим нервом импульсы идут к ядрам этого нерва. Реализуются статокинетического рефлексы с участием двигательных ядер продолговатого и среднего мозга. При угловом ускорении наблюдается медленное движение головы в сторону, противоположную направлению вращения, а затем голова быстро поворачивается в нормальное положение, чтобы впоследствии снова повторить медленное движение, и т.д. Такие движения называются нистагмом головы. Глазные яблоки реагируют на обращения аналогично - медленный поворот их в сторону, противоположную вращению, сопровождается быстрым возвращением в исходное положение (нистагм глаз). Значение нистагма состоит в том, что благодаря ему сохраняется нормальная зрительная ориентация во время движения. 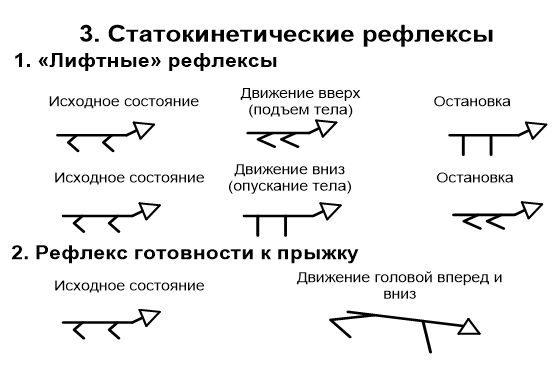 22. Характеристика нисходящих влияний ретикулярной формации на двигательные центры спинного мозга. Ретикулярная формация ствола мозга – скопление полиморфных нейронов по ходу ствола мозга. Физиологическая особенность нейронов ретикулярной формации: 1) самопроизвольная биоэлектрическая активность. Ее причины – гуморальное раздражение (повышение уровня углекислого газа, биологически активных веществ); 2) достаточно высокая возбудимость нейронов; 3) высокая чувствительность к биологически активным веществам. Ретикулярная формация имеет широкие двусторонние связи со всеми отделами нервной системы, по функциональному значению и морфологии делится на два отдела. 1) растральный (восходящий) отдел – ретикулярная формация промежуточного мозга; 2) каудальный (нисходящий) – ретикулярная формация заднего, среднего мозга, моста. Физиологическая роль ретикулярной формации – активация и торможение структур мозга. Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, т. е. тормозятся. Нисходящее влияние ретикулярной формации ствола головного мозга на спинной мозг осуществляется через ретикулоспинальный тракт. Если раздражается ретикулярная формация промежуточного мозга - преобладает тормозное влияние ретикулярной формации. Тормозное влияние происходит через вставочные тормозные нейроны (клетки Реншоу). При раздражении ретикулярной формации продолговатого мозга происходит повышение активности мотонейронов спинного мозга - нисходящее активирующее влияние. Нисходящее влияние ретикулярной формации выражено по-разному в отношении центров противоположных рефлексов. Ретикулярная формация регулирует двигательную активность совместно с мозжечком и нейронами среднего мозга (распределение тонуса). Также ретикулярная формация регулирует вегетативные функции (в составе дыхательного и сосудодвигательного центров). 23. Характеристика восходящих влияний ретикулярной формации на кору больних полушарий мозга. Ретикулярная формация ствола мозга – скопление полиморфных нейронов по ходу ствола мозга. 1) самопроизвольная биоэлектрическая активность. Ее причины – гуморальное раздражение (повышение уровня углекислого газа, биологически активных веществ); 2) достаточно высокая возбудимость нейронов; 3) высокая чувствительность к биологически активным веществам. Ретикулярная формация имеет широкие двусторонние связи со всеми отделами нервной системы, по функциональному значению и морфологии делится на два отдела: 1) растральный (восходящий) отдел – ретикулярная формация промежуточного мозга; 2) каудальный (нисходящий) – ретикулярная формация заднего, среднего мозга, моста. Физиологическая роль ретикулярной формации – активация и торможение структур мозга. Восходящие влияния ретикулярной формации направлены на регуляцию деятельности коры больших полушарий. Большинство ретикулярных волокон достигает коры через переключение в неспецифических ядрах таламуса. Действие восходящих ретикулярных влияний заключается в широкой активации корковых структур. Торможение активности ретикулярной формации ведет к наступлению сна, активация ее приводит к реакции пробуждения. Активизирующее влияние ретикулярной формации очень выражено. Через ретикулярную формацию проходит неспецифический путь передачи импульсов в кору головного мозга. Импульсы поступают к ретикулярной формации и теряют свою специфичность. От ретикулярной формации импульсы веерообразно расходятся ко всей коре головного мозга. Они стимулируют обменные процессы в коре головного мозга и повышают возбудимость нейронов коры. При удалении активирующего влияния животные засыпают. Тормозное влияние. При раздражении гигантоклеточных ядер ретикулярной формации продолговатого мозга наблюдается торможение функции коры головного мозга (сон). 24. Характеристика двигательных ядер ствола мозга и их нисходящих влияний на спинальные мотонейроны. Децеребрационная регидность и механизм ее возникновения. Вестибулярные ядра - это орган, фиксирующий изменение положения тела в пространстве и находящийся во внутреннем ухе. Возбуждение вестибулярных ядер происходит под действием адекватного раздражителя, действующего на вестибулярный аппарат. Начинающийся от ядра Дейтерса — одного из главных ядер, — а также от верхнего и медиального ядер вестибулоспинальный путь осуществляет воздействие на альфа-мотонейроны спинного мозга: нейроны вестибулярного ядра возбуждают α-мотонейроны разгибателей, причём преимущественно на осевые мышцы (мышцы позвоночного столба) и одновременно тормозят α-мотонейроны сгибателей по механизму реципрокной иннервации. При экспериментальной перерезке вестибулоспинального тракта наблюдается преобладание тонуса в мышцах-сгибателях. Также, от вестибулярных ядер продолговатого мозга идёт путь к так называемому медиальному пучку, направленному в сторону спинного мозга. Этот пучок выполняет важную функцию: соединяет воедино все ядра нервов, участвующих в регуляции активности мышц глазного яблока. Сигналы, идущие от вестибулярных ядер, попадают на продольный медиальный пучок, благодаря чему при активизации вестибулярного аппарата возникает явление нистагма. Таким образом, при раздражении вестибулярного аппарата происходит перераспределение мышечного тонуса и изменение активности мышц глазного яблока, в результате чего животное способно удержать равновесие и направить взор в нужную сторону. Красное ядро расположено в области среднего мозга. Нейроны этого ядра получают информацию от коры головного мозга и мозжечка, то есть всю информацию о положении тела в пространстве, о состоянии мышечной системы, кожи. Влияние на альфа-мотонейроны спинного мозга осуществляется с помощью руброспинального тракта. Руброспинальный тракт начинается от клеток красного ядра, расположенного в коре ножек мозга. Активация нейронов красного ядра вызывает возбуждающий постсинаптический потенциал в мотонейронах мышц-сгибателей, а в мотонейронах разгибателей — тормозные постсинаптические потенциалы. В этом отношении руброспинальный тракт сходен с кортикоспинальным трактом. Децеребрационная ригидность – резкое повышение тонуса мышц-разгибателей и относительное расслабление мышц-сгибателей, возникающие в результате перерезки стволовой части головного мозга — децеребрации. При Д. р. утрачиваются рефлексы, сохраняющие равновесие тела и его способность к движению: туловище и все конечности животного разгибаются и судорожно вытягиваются, голова запрокидывается (т.н. опистотонус). Причина Д. р.: высвобождение тонических центров продолговатого и спинного мозга из-под сдерживающего контроля ретикулярной формации продолговатого и среднего мозга. Механизм децеребрационной ригидности заключается в том, что нарушается регулирующее (тормозное) влияние красного ядра и ретикулярной формации на мотонеройны и γ-нейроны спинного мозга. Расторможенные γ-нейрoны получают раздражение от проприорецепторов мышцы. Тонус мышц-разгибателей, проприорецепторы которых раздражаются при их натяжении, стоянии или другом положении тела в пространстве, резко повышается. 25. Схема влияния приема Ендрассика на сухожильные рефлексы спинного мозга. Прием Ендрассика: обследуемому предлагают сцепить пальцы обеих рук и силой тянуть кисти в стороны. Одновременно проводят отрывистые удары молоточком по связке коленной чашечки. Обычно на этом фоне рефлекторные сокращения четырёхглавой мышцы бедра, вызывающие разгибание в коленном суставе, происходят с укорочением латентного периода ответа и увеличением его силы. Активность нейронов шейного утолщения спинного мозга или какого-то более рострально расположенного центра, возникающая при приёме Ендрассика, передается к альфа-мотонейронам спинного мозга и вызывает возбуждение тех мотонейронов, которые в отсутствие приёма Ендрассика находятся в состоянии «подпорогового возбуждения». Усиливающее воздействие приёма Ендрассика на моносинаптические рефлексы у человека изучали путём сравнения H- и T-рефлексов солеуса и блокирования активности нервных волокон. Метод Ендрассика одинаково усиливает проявления H- и T-рефлексов при условии, что они слабо работают. Н-рефлекс остаётся усиленным и в том случае, когда активность мышечных веретён в солеусе не изменяется, или когда афферентные нервные волокна типа 1α, идущие из солеуса, заблокированы. Из этого следует, что усиливающий эффект при приёме Ендрассика влияет на α-мотонейроны преимущественно не через γ-петлю, а через ослабление пресинаптического ингибирования или другой центральный механизм. |