Коллоквиум по физиологии. Колоквиум 2 ЦНС. Коллоквиум по разделу Общая и частная физиология цнс
 Скачать 1.11 Mb. Скачать 1.11 Mb.
|

|
Коллоквиум по разделу: Общая и частная физиология ЦНС Физиологические свойства нейронов и характеристика их функциональных состояний. Нейрон - структурно-функциональная единица нервной системы; электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. В зависимости от выполняемой функции нейроны делятся на 3 основные группы (По положению в рефлекторной дуге) 1.афферентные (чувствительные) 2.эфферентные (двигательные) 3.вставочные (контактные). Афферентные нейроны обеспечивают восприятие раздражения и передачу информации в ЦНС. Афферентные нейроны (чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания. Эфферентные нейроны обеспечивают передачу информации от ЦНС на периферию. Эфферентные нейроны (эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные. Вставочные нейроны обеспечивают передачу информации внутри ЦНС (с афферентных нейронов на эфферентные). Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами В зависимости от эффекта вставочные нейроны подразделяются на (по функциональному состоянию) 1. возбуждающие – оказывают возбуждающее влияние на эфферентные нейроны 2. тормозные – оказывают тормозное влияние на эфферентные нейроны. В зависимости от вида медиатора в синапсе нейрона различают: 1.холинергические нейроны (медиатор – ацетилхолин) 2.адренергические нейроны (медиаторы – адреналин и норадреналин) Физиологические свойства нейрона: возбудимость; проводимость; лабильность – это скорость физико-химических реакций лежащих в основе возбуждения (нервная ткань > лабильна, чем мышечная ткань)%; наличие постоянного тонуса, т.е. клетка находится всегда в определенном состоянии возбуждения; Рецепторная; Хранение информации; Транспортная; Синтецирующая; Кодирующая; Секреторная. Функциональные состояния нейрона. Нейрон может находиться в трех основных состояниях – 1) в состоянии покоя, 2) в состоянии активности, или возбуждения, 3) в состоянии торможения. В состоянии покоя нейрон имеет стабильный уровень мембранного потенциала. В любой момент нейрон готов возбудиться, т.е. генерировать потенциал действия, либо перейти в состояние торможения. В состоянии активности, т.е. при возбуждении нейрон генерирует а) потенциал действия б) чаще – группу потенциалов действия (серия ПД, пачка ПД, вспышка возбуждения). Частота следования потенциалов действия внутри данной серии ПД, длительность этой серии, а также скважность (интервалы) между последовательными сериями – все эти показатели широко варьируют и являются составляющей кода нейронов. Тормозные нейроны снижает свою фоновую частоту разрядов в ответ на внешний сигнал, а возбуждающиеся – увеличивают частоту фоновой активности 2. Особенности строения нейрона. Механизм развития возбуждения в нервной клетке.  I — сенсорный нейрон: 1 — окончания нейрона; 2 — аксон; 3 — ядро; 4 — тело клетки; 5— дендрит; 6 — миелиновая оболочка; 7—рецептор; 8 — орган; 9— неврилемма; II— двигательный нейрон: 1 — дендриты; 2— аксон; 3 — концевая бляшка; 4 — перехват Ранвье; 5 — ядро шванновской клетки; 6 — шванновская клетка; III — вставочный нейрон: 1 — аксон; 2 — дендриты; 3 — ядро; 4 — тело клетки; 5 — дендрон. 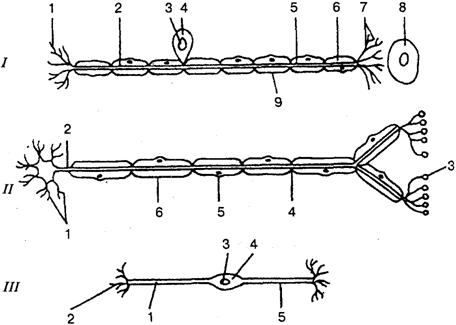 Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также специфические образования - нейрофибриллы. Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу. Дендриты — короткие и сильно разветвлённые отростки нейрона, воспринимающие раздражение или нервные импульсы от других нейронов и передающие возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами. Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии. Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной. Механизм возбуждения в нервной клетке. При возбуждении нейронов потребление кислорода возрастает. Источником энергии служит в основном глюкоза крови, собственных небольших запасов гликогена хватает лишь на 3-5 мин работы нейрона. Передача сигнала в химических синапсах ЦНС подобна таковой в нервно-мышечном синапсе, однако имеет ряд отличительных особенностей. Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных импульсов и их взаимодействие. Одна везикула (квант медиатора) содержит 1—10 тыс. молекул медиатора. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 200—300 квантов медиатора; при этом возникает небольшой ВПСП — около 0,05 мВ (миниатюрный ВПСП). (На нейронах одновременно присутствуют возбуждающие и тормозные синапсы). Пороговый потенциал нейрона равен 5—10 мВ. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль Са2+, причем количество медиатора прямо пропорционально входу ионов Са2+, при этом четыре иона Са2+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени по входам в результате пространственной суммации ВПСП возникает ПД нейрона-мишени. 1) На нейрон пришло одинаковое число ВПСП и ТПСП -> нейрон в состоянии покоя. 2) ТПСП>ВПСП – при суммации на мем-не нейрона не генерируется ПД и нейрон оказ-ся в состоянии торможения 3) ВПСП>ТПСП – деполяризация на мембране -> генерация ПД ВПСП возникает вследствие суммарного тока в клетку и из клетки через ионные каналы различных ионов согласно электрохимическому градиенту. Поступивший в пресинаптическое окончание Са2+ удаляется в среду с помощью Са-насоса. Прекращение действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично — с помощью разрушения специальными ферментами. Место возникновения ВПСП, вызывающих ПД нейрона, — тело нейрона, поскольку постсинаптические мембраны этих синапсов располагаются в непосредственной близости от аксонного холмика, где начинается развитие ПД. Близость ВПСП, возникающих в теле нейрона, к аксонному холмику обеспечивает их участие в механизмах генерации ПД. Формирование ПД начинается на мембране аксонного холмика. Синапсы на нем отсутствуют, возбудимость мембраны аксонного холмика в 3—4 раза превосходит возбудимость мембраны тела нейрона, что объясняется более высокой (примерно в 7 раз) концентрацией Na- каналов на аксонном холмике (на нем отсутствует миелиновая оболочка; порог раздражения - 10мВ, по сравнению телом – 30мВ). ВПСП, возникающие в любых участках сомы нейрона суммируются, их суммарное электрическое поле достигает аксонного холмика и вызывает его деполяризацию до КП, в результате чего начинает развиваться ПД на аксонном холмике (генераторный потенциал). Электрическое поле начального возбуждения аксонного холмика обеспечивает деполяризацию тела нейрона до КП (соматодендритный потенциал), в результате чего возникает ПД нейрона, который проводится по аксону к другой клетке (аксонный потенциал). На восходящей части ПД нейрона регистрируется перегиб на уровне 30— 40 мВ (задержка в развитии ПД), вследствие того что возбудимость мембраны тела нейрона в 3—4 раза ниже возбудимости мембраны аксонного холмика, что также является подтверждением описанного механизма формирования ПД нейрона чувствительности постсинаптической мембраны к действующему медиатору. 3. Виды межнейрональных синапсов, характеристика их свойств. Межнейрональные синапсы - это коммуникационные соединения между нейронами. По их расположению различают: Аксосоматические (когда аксоны одного нейрона оканчиваются на теле другого нейрона) аксодендритические (аксоны одного нейрона оканчиваются на дендритах другого нейрона) аксоаксональные (аксоны одного нейрона заканчиваются на аксонах другого нейрона, обычно тормозя функцию последнего) дендросоматические По способу передачи импульса различают химические синапсы и электрические синапсы (передают импульс на другую клетку с помощью специальных биологически активных веществ – нейромедиаторов, находящихся в синаптических пузырьках) Электрические синапсы – двустороннее проведение, низкая утомляемость, высокая лабильность, короткая СЗ. Химические синапсы – одностороннее проведение, высокая утомляемость, низкая лабильность, длинная СЗ. Бывают: адренергическими (норадреналин), холинергическими (ацетилхолин), пуринэргическими, с участием АМК (ГАМК, глицин, глютам), аминергетическими (дофамин, серотонин). По анатомо-гистологическому принципу. 1. нейросекреторные. 2. нервно-мышечные. 3. межнейронные. По функциональному принципу. 1. возбуждающие. 2. тормозные. 4. Принципы распространения возбуждения в нейрональной сети. Их систематическое представление и функциональное значение. Возбуждение в ЦНС распространяется по различным конфигурациям нервных цепочек. Во всех изученных нервных сетях обнаружены: конвергенция путей, дивергенция путей и реверберация. 1) Конвергенция – это схождение нескольких нервных импульсов-путей, поступление разного типа возбуждений к одним и тем же нейронам. Конвергенция многих нервных путей к одному нейрону или центру делает его интегратором соответствующих сигналов. Его состояние в каждый момент времени определяется алгебраическим сложением массы возбуждающих и тормозных входов. Иными словами, суммой всех его ВПСП и ТПСП, поступающих на данный центр или нейрон.  Мощная конвергенция обнаруживается на мотонейронах спинного мозга, нейронах ретикулярной формации ствола мозга, неспецифических ядрах таламуса. Благодаря конвергенции происходит «сжатие», уменьшение объема информации, поступающей от рецепторов к ЦНС. 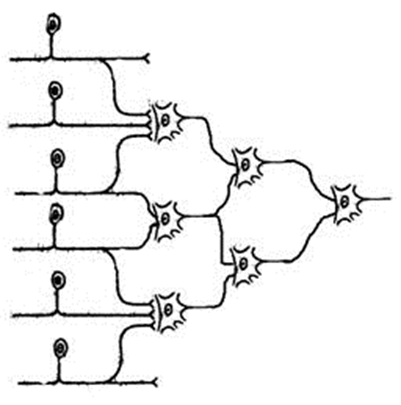 • Мультисенсорная – от нескольких сенсорных модальностей к одному нейрону. • Мультибиологический – заключается в схождении к одному нейрону возбуждений от центров, формирующих те или иные биологические состояния, для выбора мотивации (голод, жажда, боль). • Сенсорно-биологический – представляет собой комбинация двух первых, конвергирует информацию от системы и центра (схождение к нейрону импульсов от рецепторов сетчатки глаза и гипоталамического центра голода).  Дивергенция пути сигнала наблюдается у многих вставочных нейронов. Благодаря дивергенции могут формироваться процессы параллельных вычислений, что обеспечивает высокий уровень быстродействия ЦНС. Дивергенция пути обеспечивает расширение области распространения сигнала. Бывает направленная (в норме) и диффузная (у детей и при патологиях). 2) Дивергенция (иррадиация) – распространение импульсов, поступающих по афферентным волокнам от одного нейрона через коллатерали его аксона к большему числу нервных клеток. Так, существует разделение аксона чувствительного нейрона в спинном мозге на множество коллатералей. Получившиеся разветвления направляются к разным сегментам спинного мозга и в головной мозг. 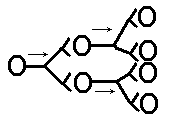 3) Реверберация - длительная циркуляция импульсов возбуждения в сетях нейронов или между различными структурами цнс. Считается, что реверберация может служить основой кратковременной памяти. 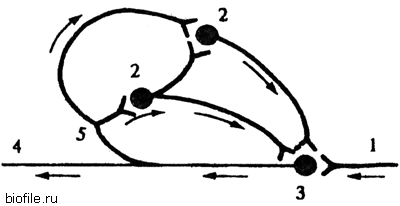 4) Мультипликация (умножение) – усиление активации нейрона за счет вовлечения в его стимуляцию дополнительных вставочных нейронов «на входах». 5) Пролонгирование – усиление активности нейрона за счет вовлечения в его стимуляцию дополнительных вставочных нейронов на его выходах. 5. Понятие нервного центра с краткой характеристикой его свойств. Нервный центр - это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т.д.). В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности. Нервные центры обладают рядом функциональных свойств и особенностей. К основным свойствам нервных центров относятся: 1) одностороннее проведение возбуждения - от рецептора к рабочему органу, обусловливается свойством синапсов односторонне проводить возбуждение. 2) Центральная задержка – время проведения возбуждения через синапсы; время анализа и синтеза поступающей в центр информации 3) В нервных центрах происходит суммация возбуждений. Суммация может быть: Пространственной – суммация постсинаптических потенциалов от последовательно приходящих импульсов через одни и те же синапсы Временной – от одновременно приходящих сигналов через разные синапсы Окклюзия – закупорка возбуждения; амплитуда ответа центра на раздражение суперпороговыми стимулами нескольких рецепторных зон меньше суммы ответов центра на раздражение каждой рецепторной зоны отдельно. Облегчение – амплитуда ответа центра на раздражение пороговыми стимулами некольких рецепторных зон больше сусммы ответа центра на раздражение каждой рецепторной зоны отдельно 4) Принцип общего конечного пути – количество «входов» в центр всегда больше «выходов». 5) Рефлекторное последействие — реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено либо длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, либо в результате циркуляции (реверберации) возбуждения в нейронной сети. 6) Нервные центры обладают высокой чувствительностью к недостатку кислорода и к химическим веществам. 7) Нервные центры, как и синапсы, обладают быстрой утомляемостью (в связи с синаптической депрессией, связанной с окончанием медиатора) в отличие от нервных волокон, которые считаются практически неутомляемыми. Они обладают низкой лабильностью. 8) В нервных центрах легко возникает процесс торможения. 9) Нервные центры обладают пластичностью — способностью изменять собственное функциональное назначение, частично восстанавливать утраченные функции. 10) Принцип фракционирования – разный уровень возбудимости определяют группы клеток центра 11) Тонус – постоянный уровень активности нейронов. Определяется величиной МПП клеток, общей суммарной активностью. Зависит от приходящих ВПСП и гуморальных факторов. 12) Принцип субординации (иерархичность) 13) Принцип доминанты – механизм повышения возбудимости центра с жизненно важной функцией путем 1) усиления проводимости; 2) торможения центров недоминантного плана (латеральное торможение). 14) Посттетаническая потенциация – ответная реакция на один и тот же раздражитель при длительной подпороговой стимуляции увеличивается. 6. Общая характеристика процесса торможения в ЦНС. Его биологическая роль. Торможение в ЦНС — это активный процесс, проявляющийся в препятствии возникновения возбуждения или распространении уже возникшего возбуждения. Торможение может наступить не только в результате непосредственного воздействия на нервные центры, но и на рецепторы. В основе – тормозные процессы на мембране нейронов: длительная поляризация, длительная деполяризация, гиперполяризация. 1) Торможение может развиваться только в форме локального ответа (т.е. оно будет затухающим; способно суммироваться; зависит от силы раздражителя) 2) Всегда следствие возбуждения 3) Требует затрат энергии Центральное торможение: Первичное торможение: возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы. Виды первичного торможения Постсинаптическое — основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Возвратное — нейрон воздействует на клетку, которая в ответ тормозит этот же нейрон. Реципрокное — это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон. Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клетками сетчатки, что создаёт условия для более четкого видения предмета. Возвратное облегчение — нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками. Пресинаптическое — возникает в обычных нейронах, связано с процессом возбуждения. |