Экология Часть 1. Курс лекций Красноярск сфу 2 010 удк 574(042. 4)(075) ббк 28. 081 я 73 Ч90 а вторы
 Скачать 1.38 Mb. Скачать 1.38 Mb.
|

|
Пустыни В зависимости от горных пород, слагающих территорию, различают: глинистые, каменистые и песчаные пустыни. Вопреки расхожему представлению о пустынях как обширных пространствах, где тянутся бесконечные волнистые ряды песчаных барханов, лишь одна пятая часть площади мировых пустынь покрыта песком. Однако и там умещается немало впечатляющих песчаных морей. В Сахаре песчаные пустыни, эрги, покрывают многие десятки тысяч квадратных километров. Песок, который смывается туда с соседних нагорий, образуется в результате выветривания пород пустыни. Он постоянно переносится ветром с места на место и в конечном счете скапливается в низинах и впадинах. Каменистые пустыни бывают нескольких типов, в зависимости от типа поверхности. Ее могут образовывать камень, щебень, галька, гипс. Поверхность одних пустынь хорошо проницаема для воды, других создает плотную водонепроницаемую корку. В первом случае вода уходит на глубину, недоступную корням растений. Во втором – испаряется с поверхности, еще больше закрепляя корку пустыни. Там, где раньше была вода, образуются соли. В некоторых местах их концентрация настолько велика, что они создают корку на поверхности. Есть места, толщина которых составляет 15 см с торосами высотой до метра. Если влага не совсем испарилась, солончаки имеют вид топкого болота. Одними из наиболее распространенных типов пустынь являются каменистые, щебнистые, щебнисто-галечниковые и гипсовые пустыни. Их объединяет грубость, твердость и плотность поверхности. Водопроницаемость каменистых грунтов различна. Наиболее крупные галечниковые и щебнистые обломки, залегающие достаточно рыхло, легко пропускают воду, и атмосферные осадки быстро просачиваются на большие глубины, недоступные растениям. Но чаще, однако, распространены поверхности, где галечник или щебень сцементированы песчаными или глинистыми частицами. В таких пустынях каменистые обломки лежат плотно, образуя так называемую пустынную мостовую. В пустынях количество осадков уменьшается до 200 мм/ год, в связи с их уменьшением и нерегулярностью выпадения уровень бесплодности (засушливости) почв повышается. Растительный покров здесь не сплошной, представлен многолетними (древесными ксерофильными) и однолетними (терофитами) растениями с очень коротким вегетативным периодом. Флора пустынь представлена большим разнообразием видов, приспособившихся к засушливым условиям. Здесь имеется целый ряд многолетних растений с постоянными воздушными органами, растущих в пустынях даже там, где среднее годовое количество осадков ниже 50 мм. Редукция листвы, ее ксероморфизм, говорит о приспособленности растений к уменьшению потерь воды при эвапотранспирации. Каменистые пустыни Сахары (гамады), занимающие до 70 % ее площади, чаще лишены высшей растительности. Лишь на отдельных каменистых осыпях закрепляются подушковидные кустарники фреодолия и лимонаструм. Более увлажненные пустыни Средней Азии, хотя и разреженно, но равномерно покрыты полынью и солянками. На песчано-галечниковых равнинах Центральной Азии распространены низкорослые заросли саксаула. Животный мир пустынь составляет множество видов пресмыкающихся и грызунов, адаптировавшихся к отсутствию влаги: воду они получают из пищи. Крупные травоядные зооценоза пустынь также приспособились к недостатку воды. Однако они не могут обходиться без воды в отличие от пресмыкающихся и грызунов. Рекорд сопротивления жажде удерживает сахарская антилопа аддакс (Addax nasomaculatus). Аддаксы питаются травами и низкорослыми кустарниками. Они преодолевают большие расстояния по пустыне в поисках редкой растительности. Они обходятся без питьевой воды большую часть своей жизни, а необходимый для выживания запас воды получают от растений, которыми питаются. На данный момент аддаксы одни из наиболее редких млекопитающих в мире – менее чем 250 диких животных. Для пустынных ландшафтов характерны небольшие значения биомассы и продуктивности (от 10 до 100 ц/га и от 5 до 50 ц/га соответственно). Прямые водные связи в этих ландшафтах значительно ослаблены, что определяет независимость друг от друга основных компонентов ландшафта, например, элювиальных почв и грунтовых вод. Продуктивность сельскохозяйственных угодий обычно несколько ниже, чем природных экосистем в той же зоне. Поля часть года пустуют, и на них обычно выращивают всего один какой-либо вид, который не в состоянии полностью использовать все имеющиеся ресурсы. Однако при интенсивном земледелии продуктивность полей может приближаться к максимальной, хотя человеку приходится вкладывать в это много дополнительных средств. Знание законов биологической продуктивности и потерь энергии в цепях питания имеет большое практическое значение. На их основе можно сознательно и грамотно строить хозяйственную деятельность таким образом, чтобы не подрывать воспроизводительные способности природных и антропогенных систем и получать возможно большую первичную и вторичную продукцию. Для человека энергетически выгоднее растительное питание, а наиболее дорого – использование в пищу хищных видов. Так, по энергии, затраченной на рост, 1 кг окуня или щуки обходится природе в cемь раз дороже, чем 1 кг говяжьего мяса. Поэтому плотоядные животные разводятся людьми в редких случаях, например в пушном звероводстве. Широкое одомашнивание нашими предками таких видов, как свиньи и куры, не случайно. Они характеризуются высоким коэффициентом использования энергии на рост, т. е. перевода пищи в собственную биомассу. Одна растительная пища, как правило, для людей недостаточно полноценна, так как подавляющее большинство растений не обеспечивает людей некоторыми незаменимыми аминокислотами, входящими в состав животных белков. Производство вторичной продукции через выращивание животных, а также добыча диких видов (в основном путем рыболовства) – очень важное условие благополучия общества. Одна из самых злободневных для современного человечества проблем – это так называемое белковое голодание, недостаток животной пищи в рационах людей во многих районах мира. Поэтому значительное внимание необходимо уделять повышению продуктивности естественных и искусственных экосистем. Пути повышения продуктивности естественных экосистем Для улучшения растительного покрова и повышения продуктивности различных экосистем особенно эффективны следующие мероприятия: внесение удобрений на луга и пастбища; орошение; осушение травяных болот, используемых под сенокос; проведение поверхностного рыхления почвы, удаление мохового покрова, срезание кочек на лугах и пастбищах; подсев кормовых трав (злаков, бобовых) и борьба с сорняками на лугах и пастбищах. Подобное вмешательство человека в жизнь естественных экосистем способствует улучшению аэрации почвы, обогащению ее питательными веществами и в конечном счете приводит к заметному повышению продуктивности экосистем. Пути повышения продуктивности искусственных экосистем Стратегическим направлением развития сельскохозяйственного производства является повышение урожайности культурных растений, что позволит обеспечить растущее население продовольствием при сохранении прежних посевных площадей. Этого можно достичь следующими путями: выведением и внедрением в производство новых высокоурожайных сортов растений; соблюдением высокой культуры земледелия, которая включает научно обоснованные приемы обработки почвы, полива и внесения оптимальных доз удобрений, соблюдение сроков посева и глубины заделки семян; созданием интегрированной системы защиты растений, которая наряду с химическими методами борьбы с вредителями, сорняками и болезнями включает правильные севообороты и применение биологических методов борьбы; орошением и др. Радикальным способом повышения продуктивности искусственных экосистем является культивирование растений в теплицах с использованием гидропоники. Индустриальная технология выращивания растений В силу ряда причин (строительство городов, промышленных предприятий, электростанций, дорог и т. д.) количество посевных площадей во всем мире неуклонно снижается. Поэтому для того чтобы обеспечить продуктами питания возрастающее население нашей планеты, необходимо развивать агроиндустрию, которая базируется на культивировании растений в искусственно созданных условиях. Агроиндустрия имеет немало преимуществ в сравнении с традиционным естественным земледелием: она дает возможность избавиться от капризов погоды, сорняков и вредителей, использовать в полной мере вносимые удобрения, создать оптимальные условия для выращивания растений в течение всего года, в том числе в осенне-зимний период, освободить значительные территории и решить многие другие сельскохозяйственные, экологические и демографические проблемы. Выращивание растений, частично или полностью изолированных от внешней среды, производится в парниках, теплицах, климатических камерах (фитокамерах, фитотронах). Наиболее широко и повсеместно используются для такой цели теплицы – строения, покрытые стеклом или прозрачной пленкой. Современные теплицы оборудованы автоматической системой поддержания микроклимата, дождевания и подкормки. Для обеспечения оптимального водно-воздушного режима под тепличным грунтом прокладывается дренажная система надпочвенного и подпочвенного обогрева. По сути, почва является только поставщиком минеральных элементов и механической опорой растений. Эти функции с еще большим успехом могут взять на себя созданные специалистами искусственные почвозаменители. Более того, в последнее время получил широкое развитие беспочвенный способ выращивания растений. 7.3. Изменения продуктивности и биомассы в ходе смены (сукцессии) экосистем Динамика экосистемы определяется серией сменяющих друг друга сообществ. Сукцессия (от лат. succesio – преемственность, наследование) – последовательная смена во времени одних биогеоценозов другими на определенном участке земной поверхности. Сукцессионный ряд – цепь сменяющих друг друга биоценозов. Процессы сукцессии занимают определенные промежутки времени. Чаще всего это годы и десятки лет, но встречаются и очень быстрые смены сообществ, например, во временных водоемах, и очень медленные – вековые изменения экосистем, связанные с эволюцией на Земле. Сукцессия завершается формированием сообщества, наиболее адаптированного по отношению к комплексу сложившихся климатических условий. Такое сообщество было названо Ф. Клементсом климаксформацией или просто климаксом, хотя в современной литературе по экологии иногда встречается и другой термин-синоним – зрелое сообщество. Концепция «климакса» подразумевает, что в пределах региона с более или менее однородным климатом фитоценозы, завершившие сукцессионный процесс, образуют климаксные сообщества независимо от того, с какого типа начиналась сукцессия. Например, в верховьях р. Большая Кеть (Томская область) климаксовыми сообществами являются кедрачи, ельники, пихтарники в различном сочетании на супесчаных, суглинистых, глинистых почвах и сосняки на песчаных почвах. Климаксовые сообщества могут поддерживать наиболее благоприятные для себя локальные условия окружающей среды в устойчивом состоянии, компенсируя любые возникающие случайные возмущения, сохраняя равновесие между биотическими и абиотическими компонентами до следующего нарушения. Причиной начала процесса сукцессии в ряде случаев являются изменения фундаментальных свойств среды обитания, возникающие под влиянием комплекса факторов. Такие факторы бывают естественными – отступление ледников, наводнения, землетрясения, извержения вулканов, пожары, а также антропогенными – расчистка лесных угодий, распашка участков степи, открытая добыча полезных ископаемых, создание прудов и водохранилищ, пожары, загрязнение экологических систем. В зависимости от обстоятельств, предшествовавших началу процесса, сукцессии подразделяют на следующие: антропогенные, вызванные хозяйственной деятельностью человека; катастрофические, связанные с какими-либо катастрофическими для экосистемы природными или антропогенными факторами (извержение вулкана, землетрясение, разливы нефти на большой территории); пирогенные, вызванные пожаром, независимо от его причин; зоогенные (фитогенные), вызванные необычно сильным воздействием животных (растительности), как правило, в результате их массового размножения (завоза чуждых видов человеком). По общему характеру сукцессии делят на первичные и вторичные. Первичные сукцессии. Они начинаются на субстрате, не измененном (или почти не измененном) деятельностью живых организмов. Так, через серию промежуточных сообществ формируются устойчивые сообщества на скалах, песках, обрывах, остывшей вулканической лаве, глинах после отступления ледника или прохождения селя и т. п. Одна из основных функций сукцессии такого рода – постепенное накопление органических остатков и, как результат, создание почвы. Первичная сукцессия от голой скальной породы к зрелому лесу может занять от нескольких сотен до тысяч лет. Вторичные сукцессии. Они развиваются на субстрате, первоначально измененном деятельностью комплекса живых организмов, существовавших на данном месте ранее — до пожара, наводнения, вырубки и т. п. В таких местах обычно почва или донные отложения не уничтожены, т. е. сохраняются богатые жизненные ресурсы, и сукцессии чаще всего бывают восстановительными. Здесь в почве могут сохраняться семена, споры и органы вегетативного размножения, например корневища, которые будут влиять на сукцессию. Существует множество классификаций сукцессий по показателям, которые могут меняться в ходе сукцессии или по причинам смен: по масштабу времени (быстрые, средние, медленные, очень медленные), по обратимости (обратимые и необратимые), по степени постоянства процесса (постоянные и непостоянные), по происхождению (первичные и вторичные), по тенденциям изменения продуктивности (прогрессивные и регрессивные), по тенденции изменения видового богатства (прогрессивные и регрессивные), по антропогенности (антропогенные и природные), по характеру происходящих во время сукцессии изменений (автотрофные и гетеротрофные). В зависимости от целей исследователя подобные классификации можно строить на любом логическом основании, а число их можно увеличивать до бесконечности. Если классифицировать сукцессии на основе протекающих процессов, то можно выделить две основные группы: эндогенные, происходящие в результате функционирования сообществ, и экзогенные, происходящие в результате внешнего воздействия. Движущей силой эндогенных сукцессий является несбалансированный обмен сообществ. На рис. 12 представлена зоогенная сукцессия, начавшаяся после вспышки массового размножения сибирского шелкопряда в темнохвойных лесах Томской области.  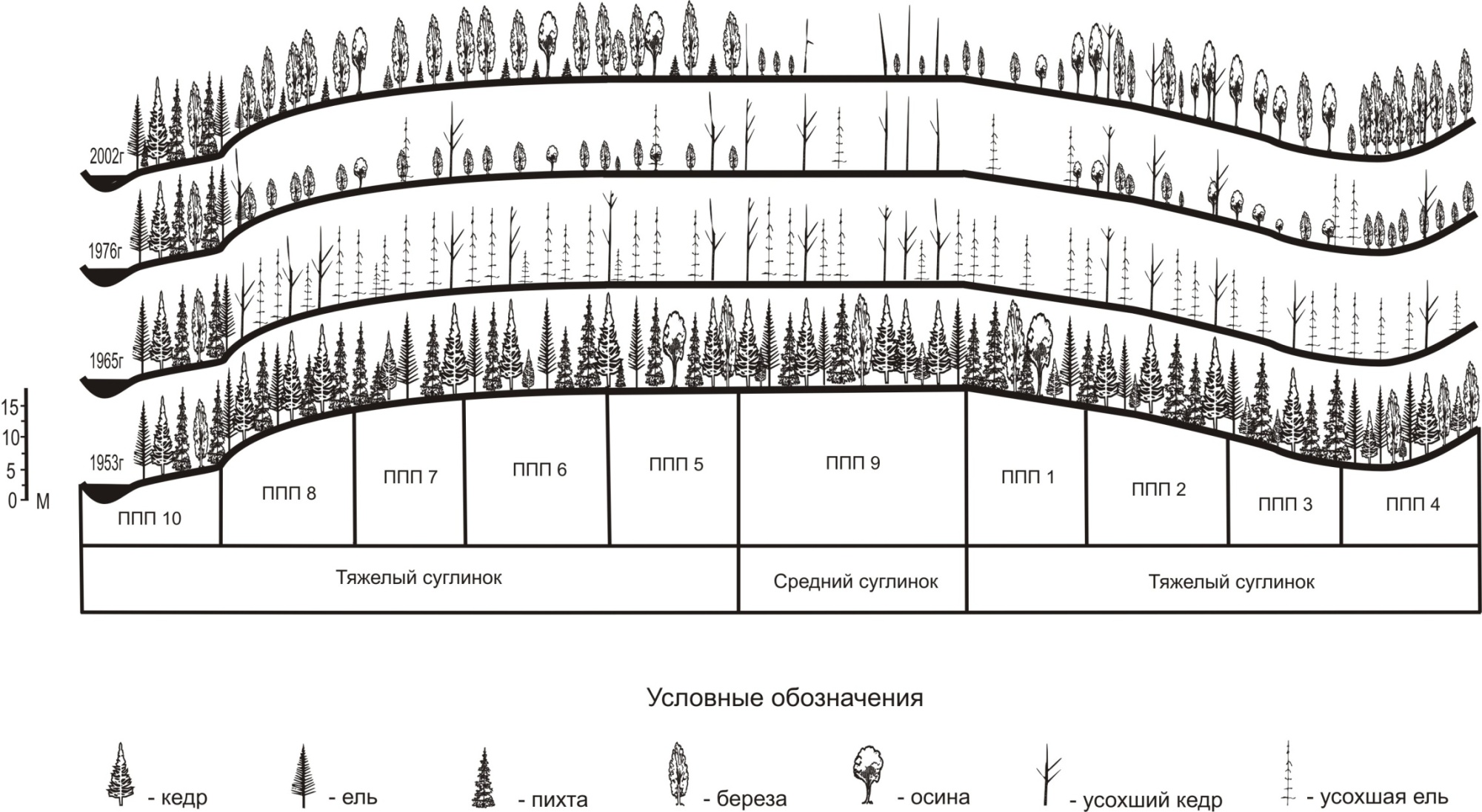 Р  ис. 12. Динамика изменения растительности после вспышки массового размножения сибирского шелкопряда ис. 12. Динамика изменения растительности после вспышки массового размножения сибирского шелкопряда Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса: 1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы, но по мере развития сукцессии эти показатели возрастают. 2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания. 3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. 4. Интенсифицируются процессы круговорота веществ, поток энергии и дыхание экосистем. 5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительные сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах. 6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обусловливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического плана. 7. В зрелой стадии климаксного сообщества биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса. Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а, во- вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов. Литература к главе 1
18. http://www.wood.ru 19. http://www.edpscirnces.org/radiopro 20. http:/www.ecology-portal.ru 21. http:/www/oeco.ru |