Экология Часть 1. Курс лекций Красноярск сфу 2 010 удк 574(042. 4)(075) ббк 28. 081 я 73 Ч90 а вторы
 Скачать 1.38 Mb. Скачать 1.38 Mb.
|

|
5.2. Основные характеристики популяции Численность и плотность популяций. Численность популяции – это общее количество особей на данной территории или в данном объеме, зависит от соотношения интенсивности размножения (плодовитости) и смертности. Плотность популяции определяется количеством особей или биомассой на единицу площади либо объема. Плотность популяции зависит от трофического уровня, на котором находится популяция. Чем ниже трофический уровень, тем выше плотность; чем крупнее животные внутри уровня, тем больше их биомасса. 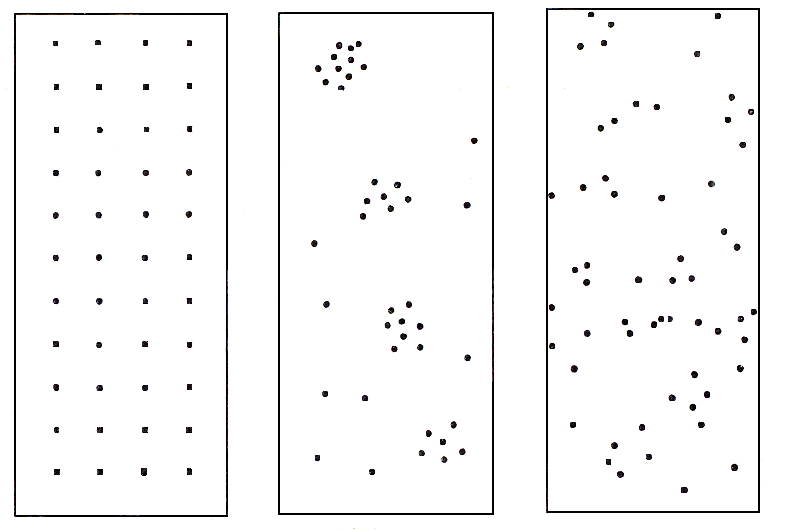 а б в Рис. 3. Три основные типа распределения особей или групп особей в популяции: а – равномерное распределение; б – групповое распределение; в – случайное распределение (по Р. Дажо,1975) Особи, составляющие популяции, имеют различные типы пространственного распределения, выражающие их реакции на различные влияния, например, добычу и благоприятные физические условия или конкурентные реакции. Различают три типа распределения или расселения особей внутри популяции: равномерное (редко, при острой конкуренции – некоторые хищные рыбы), случайное (только в однородной среде), групповое (наиболее часто) (рис. 3). В любом из них имеются сгущения, или центры плотности, между которыми население разрежено или отсутствует. Постоянные устойчивые центры плотности населения вида и представляют собой, как правило, популяции. Знание типа распределения организмов имеет большое значение при оценке плотности популяций методом выборки при экологических исследованиях. Рождаемость и смертность. Динамика численности популяций находится в тесной зависимости от рождаемости или плодовитости и смертности. Рождаемость – это способность популяции к увеличению численности, характеризует частоту появления новых особей в популяции. Различают рождаемость абсолютную и удельную. Абсолютная (общая) рождаемость – число новых особей (Nn), появившихся за единицу времени (t). Удельная рождаемость выражается в числе новых особей на особь в единицу времени (для человека – число детей в год на 1 000 человек). В популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей для данного вида. Оно достигается лишь в идеальных условиях при отсутствии лимитирующих факторов. Обычно же существует экологическая, или реализуемая, рождаемость, возникающая в обычных или специфичных условиях среды, выработанная исторически как приспособление, которое обеспечивает пополнение убыли в популяции. Смертность – это количество особей, погибших в популяции за определенный период. Абсолютная (общая) смертность – это число особей (Nm), погибших в единицу времени (t). Удельная смертность (d) выражается отношением абсолютной смертности к численности популяции. Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д. Как и рождаемость, смертность, особенно у высших организмов, широко варьирует с возрастом. Полная картина смертности в популяции последовательно описывается статистическими таблицами выживания,специально разработанными в демографических исследованиях. На основе таблиц выживания строятся кривые выживания. Такие кривые, построенные в полулогарифмическом масштабе, причем интервалы времени по оси абсцисс отложены как процент средней продолжительности жизни (или как процент общей продолжительности жизни), позволяют сравнивать виды с очень разной продолжительностью жизни. Кривые выживания подразделяют на три общих типа, показанных на рис. 4. 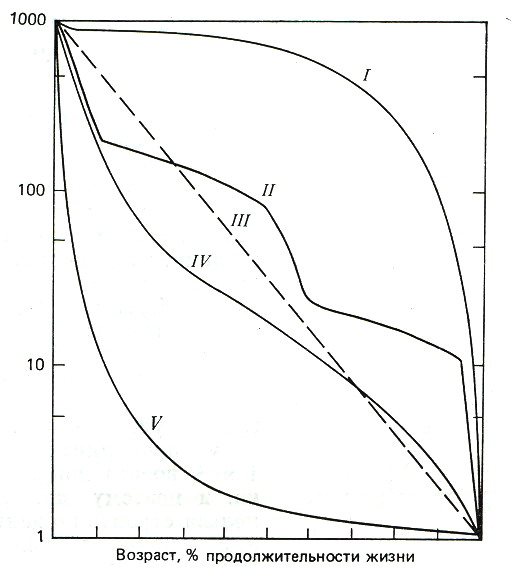 Рис. 4. Кривые выживания: I – выпуклая кривая; II – ступенчатая кривая; III – теоретическая кривая; IV – сигмоидная кривая, приближающаяся к Ш; V – вогнутая кривая (по Ю. Одуму, 1986) По оси ординат (см. рис. 4) отложено число выживших на тысячу особей (логарифмическая шкала), по оси абсцисс – возраст в процентах от продолжительности жизни. 5.3. Структура популяции Возрастная и половая структура. Популяция состоит из разных по возрасту и полу особей. На соотношение их влияют общая продолжительность жизни, время достижения половой зрелости, интенсивность размножения – особенности, вырабатываемые в процессе эволюции, как приспособления к определенным условиям. По отношению к популяции обычно выделяют три экологических возраста: пререпродуктивный, репродуктивный, пострепродуктивный. Следствием правила максимальной плодовитости (рождаемости) популяции служит правило стабильности возрастной структуры: любая естественная популяция стремится к стабильной возрастной структуре, четкому количественному распределению особей по возрастам. Это правило сформулировано А. Лоткой в 1925 г. и приложимо лишь к высшим организмам с возрастной структурой популяции. Это правило дополняется правилом стабильности соотношения полов и в совокупности они составляют правило стабильности половозрастной структуры популяции. Для описания возрастной структуры в популяции выделяют возраст-ные группы, состоящие из организмов одного возраста, и оценивают численность каждой из этих групп. Результаты представляют в виде диаграмм или пирамиды (рис. 5). 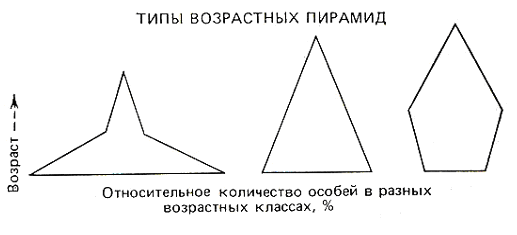 а б в Рис. 5. Возрастные пирамиды: а – популяции с высокой численностью, %, молодых особей; б – популяции с умеренной численностью, %, молодых особей; в – популяции с малой численностью, %, молодых особей (по Ю. Одуму, 1986) Генетический механизм определения пола обеспечивает расщепление потомства по полу в отношении 1:1. Но в силу разной жизнеспособности мужского и женского организмов это первичное соотношение нередко отличается от вторичного (при рождении) и третичного (для взрослых особей). У человека вторичное соотношение полов – 100 девочек на 106 мальчиков, к 16–18 годам выравнивается и к 50 годам может составлять 100 к 85. Для разных видов соотношение полов может очень различаться, и у некоторых видов пол определяется и генетическими, и экологическими факторами. Экологическая структура популяции – не только подразделенность по полу и возрасту, но и на группы особей, находящихся в специфических связях с окружающей средой. Иными словами, экологическая структура – различия между особями по экологическим свойствам в популяции. Так, известно, что пищевые спектры самцов и самок особей разных возрастных групп различны, поэтому можно говорить о трофическом полиморфизме особей в популяции. Например, исследование биологии якутского подвида соболя показало, что осенью самцы и самки питаются полевками, более крупными млекопитающими и птицами, но при этом самцы соболя чаще используют в пищу рыбу, а самки – мышевидных грызунов. В конце зимы у самок соболя рацион становится более разнообразным, чем у самцов. Кроме того, избегание хищников особями зависит от возраста, опыта, здоровья особей. В популяции можно выделять группировки особей по особенностям двигательной активности и, как следствие, по интенсивности использования территории. В популяции всегда есть различия между группами особей и по фенологии, т. е. по срокам выхода, например, из спячки, диапаузы и т. д., вплоть до появления сезонных рас у растений. Таким образом, структурированность – непременная черта любой популяции. Любое усложнение экологической структуры популяции является адаптацией, позволяет более дифференцированно использовать природную среду. Кратко рассмотренные структурные характеристики популяции не статичны, а изменяются в процессе ее существования. Изменения касаются и величины ареала популяции, и численности особей ее составляющих. 5.4. Законы изменения численности популяции В природе нет ни одного вида организмов, в популяциях которого численность особей из поколения в поколение оставалась бы постоянной. Во всех популяциях происходят изменения численности, резкие в одних случаях или менее заметные в других. Несомненно, существуют верхние и нижние пределы численности популяции, выше и ниже которых популяция перестает существовать как генетическая интегрированная система. Рассматривая вопрос об оптимальных размерах популяции в данной среде, следует учитывать поддерживающую емкость или кормовую продуктивность среды. Чем выше поддерживающая емкость (К), тем больше максимальный размер популяции, который может существовать неопределенно долгое время в данном местообитании. Дальнейшему росту популяции будут препятствовать один или несколько лимитирующих факторов. В связи с тем, что любая популяция обладает определенной генетической, фенотипической, половозрастной и другой структурой, она не может состоять из меньшего числа индивидов, чем необходимо для обеспечения стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды. В этом состоит принцип минимального размера популяций. Ю. Одум (1986) сформулировал правило популяционного максимума. Популяции эволюционируют так, что регуляция их плотности осуществляется на значительно более низкой по сравнению с верхней «асимптотой» емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы энергии и пространства. Рост популяций и кривые роста. Если рождаемость в популяции превышает смертность, то популяция, как правило, будет расти. В принципе любая популяция способна экспоненциально увеличивать свою численность. Экспоненциальная модель роста численности популяций представлена на рис. 6. 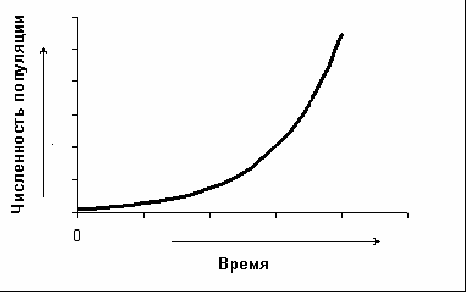 Рис. 6. Экспоненциальный (J-образный) рост популяции (jpg – element.ru) Уравнение экспоненциального роста имеет вид Nt = N0 ert,где Nt – численность популяции в момент времени t;N0– численность популяции в начальный момент t0; е – основание натуральных логарифмов (2,7182); r – удельная скорость роста. Для того чтобы экспоненциальный рост численности продолжался в течение некоторого времени, необходимо только одно условие: постоянное значение показателя r. В том случае, когда величины численности приводятся в логарифмическом масштабе, график экспоненциального роста приобретает вид прямой линии. Поэтому иногда экспоненциальный рост называют логарифмическим. Уравнение экспоненциального роста в логарифмической форме имеет следующий вид: lnNt = lnNo+rt, т. е., по сути, это просто уравнение прямой, причем коэффициент rхарактеризует угол наклона ее к осям. Если, например, какая-нибудь бактерия в благоприятной питательной среде делится каждые 20 мин, то при сохранении таких темпов деления потомство одной бактериальной клетки через 36 ч даст массу, которая покроет весь земной шар сплошным слоем толщиной 30 см, а еще через 2 ч толщина этого слоя достигнет 2 м. На самом деле в природе экспоненциальный рост популяций организмов или не происходит вообще, или же происходит, но в течение очень непродолжительного времени, сменяясь затем спадом численности или выходом ее на стационарный уровень. В принципе возможны несколько вариантов прекращения экспоненциального роста численности. Первый вариант – это чередование периодов экспоненциального роста численности с периодами резкого (катастрофического) спада, вплоть до очень низких значений. Подобная регуляция (а под регуляцией численности понимается действие любых механизмов, приводящих к ограничению роста популяции) наиболее вероятна у организмов с коротким жизненным циклом, обитающих в местах с резко выраженными колебаниями основных лимитирующих факторов, например у насекомых, живущих в высоких широтах. Очевидно также то, что такие организмы должны иметь покоящиеся стадии, позволяющие им пережить неблагоприятные сезоны. Второй вариант – это резкая остановка экспоненциального роста и поддержание популяции на постоянном (стационарном) уровне, вокруг которого возможны различные флуктуации. Третий вариант – это плавный выход на плато. Получающаяся при этом S-образная форма кривой указывает на то, что по мере увеличения численности популяции скорость роста ее не остается постоянной, а снижается. S-образный рост популяций наблюдается очень часто как в лабораторных экспериментах, так и при вселении видов в новые местообитания (рис. 7). 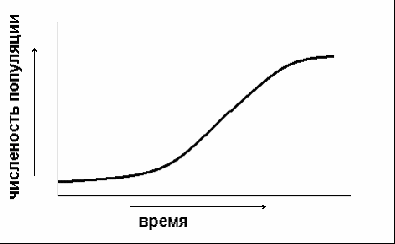 Время  Рис. 7. S-образная кривая роста популяции (gif - res.tsu.ru/zadachn/Image9.gif) Для описания S-образного «роста» может быть использовано множество различных уравнений, но наибольшую популярность получило самое простое из них — так называемое логистическое. Логистическое уравнение нагляднее всего записать в дифференциальной форме: где r – удельная скорость роста; N – текущая численность популяции; K = N – максимально возможная численность популяции. Для соблюдения экспоненциального роста необходимо, чтобы показатель rбыл постоянной величиной (r = const), для осуществления же логистического роста необходимо, чтобы показатель rснижался по линейному закону при увеличении численности популяции N. Логистическая кривая не раз использовалась и при описании результатов лабораторных опытов по культивированию тех или иных мелких организмов в ограниченном пространстве при ограниченном поступлении пищевых ресурсов. Такие зависимости в 1920–1940-е гг. были получены для бактерий, дрожжей, простейших, мелких ракообразных и ряда насекомых. Колебания численности и гомеостаз популяций. В природе численность популяций испытывает колебания. Численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции (теория лимитов популяционной численности Х. Г. Андреварты-Л. К. Бирча – 1954). Поддержание определенной численности, или равновесное состояние, получило название гомеостаза популяций. Регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида (К. Фридерикс, теория биоценотической регуляции численности популяции – 1927). В естественных условиях вероятность того, что все условия окажутся благоприятными для популяции очень низка. Как правило, один или несколько абиотических и биотических факторов становятся лимитирующими. Сочетание данных лимитирующих факторов называют сопротивлением среды. Совокупность всех факторов, способствующих увеличению численности популяции, называется биотическим потенциалом. Рост, снижение или постоянство численности популяции зависят от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей). Обеспечение популяционного гомеостаза лежит в основе устойчивого функционирования всего ряда взаимосвязанных систем повышающейся интегрированности. При всем многообразии механизмов популяционного гомеостаза их можно сгруппировать в три важнейшие функциональные категории: 1 – поддержание адаптивного характера пространственной структуры; 2 – поддержание генетической структуры; 3 – регуляция плотности населения. Отсюда принцип изменения популяций можно сформулировать следующим образом: Изменение популяции какого-либо вида – это результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей среды. Данное равновесие называют динамическим или непрерывно регулирующимся, так как факторы сопротивления среды редко остаются неизменными в течение длительного времени. Подобные циклические колебания обычно продолжаются неопределенно долго. Это объясняет обобщающее правило максимума размера колебаний плотности популяционного населения: «Существуют определенные верхние и нижние пределы для средних размеров популяций, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени». Правило максимума размера колебаний плотности популяций можно назвать законом количественной константности популяционного населения. Отклонение от этого закона или правила свидетельствует о неблагополучной ситуации, является биоиндикатором разлада в экосистеме. Периодические изменения численности популяций – одна из важнейших характеристик популяции как элементарной эволюционной единицы. Основное эволюционное значение популяционных волн состоит в том, что они приводят к резкому колебанию концентраций разных генотипов в популяции, как бы подставляя оказавшиеся в заметной концентрации генотипы под действие естественного отбора. 5.5. Экологические стратегии популяций Приспособления организмов в конечном счете направлены на повышение вероятности выжить и оставить потомство. Экологическая стратегия популяции – это ее общая характеристика и от особенностей вида, и от условий среды. Экологические стратегии разнообразны. Однако среди этого многообразия можно выделить два крайних типа, которые называются r-стратегией и К-стратегией: r-стратегия – определяется отбором, направленным прежде всего на повышение скорости роста популяции и, следовательно, таких качеств, как высокая плодовитость, ранняя половозрелость, короткий жизненный цикл, способность быстро распространяться на новые местообитания и пережить неблагоприятное время в покоящейся стадии; К-стратегия – направлена на повышение выживаемости в условиях уже стабилизировавшейся численности. Это отбор на конкурентоспособность, повышение защищенности от хищников и паразитов, повышение вероятности выживания каждого потомка. Эти две стратегии по существу представляют два различных решения одной задачи – длительного выживания вида. Виды с r-стратегией быстрее заселяют нарушенные местообитания (обнаженная горная порода, лесные вырубки), чем виды с К-стратегией, так как они легче распространяются и быстрее размножаются. Виды с К-стратегией более конкурентоспособны и они обычно вытесняют r-виды, которые тем временем перемещаются в другие нарушенные местообитания. Высокий репродуктивный потенциал r-видов свидетельствует, что, оставшись в каком-либо местообитании, они быстро использовали бы допустимые ресурсы и превысили поддерживающую емкость среды, а затем популяция погибла бы. Виды с r-стратегией занимают данное местообитание в течение жизни одного или, самое большее, нескольких поколений. В дальнейшем они переселяются на новое место. Отдельные популяции могут регулярно вымирать, но вид при этом перемещается и выживает. В целом эту стратегию можно охарактеризовать как стратегию «борьбы и бегства». Следует отметить, что одну и ту же среду обитания разные популяции могут использовать по-разному, поэтому в одном и том же местообитании могут сосуществовать виды с r- и К-стратегией. Между этими крайними стратегиями существуют переходы. Ни один из видов не подвержен только r- или только К-отбору. В целом же r- и К-стратегии объясняют связь между разнокачественными характеристиками популяции и условиями среды. |