Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

5.3. Классификация синапсов ЦНС, медиаторы синапсов цнс и их функциональное значение. Свойства синапсов ЦНС.Синапсами называются контакты нейронов с другими нервными клетками или рабочими органами. Аксон каждого нейрона, подходя к другим нервным клеткам, ветвится и образует многочисленные окончания на телах, дендритах и аксонах этих клеток. Так, на теле мотонейрона может быть около 3500 (в ретикулярной формации до 40 000 синапсов. Одно нервное волокно может образовать до 10 000 синапсов на телах многих нервных клеток. Классификация синапсов. Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала. По местоположению выделяют нервно-мышечные и нейро-нейрональные синапсы, последние в свою очередь делятся на аксо-соматические, аксо-аксональные, аксо-дендритические, дендро-соматические. По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими. По способу передачи сигнала синапсы делятся на электрические, химические, смешанные. Структура и механизм передачи возбуждения в химических синапсах. Структурно такие контакты представлены пресинаптической частью, синаптической щелью и постсинаптической частью. Пресинаптическая часть химического синапса образуется расширением аксона по его ходу или окончания. В пресинаптической части имеются агранулярные и гранулярные пузырьки. Пузырьки (кванты) содержат медиатор. Ферменты, необходимые для образования нейромедиаторов, синтезируются в перикарионе и транспортируются к синаптической терминали по аксонам. В пресинаптическом расширении находятся митохондрии, обеспечивающие синтез медиатора, гранулы гликогена и др. При многократном раздражении пресинаптического окончания запасы медиатора в синаптических пузырьках истощаются. Пузырьки бывают мелкие (диаметр порядка 50 нм) и крупные (диаметр 100–200 нм). Мелкие синаптические пузырьки содержат «классические» медиаторы (см. ниже). Крупные везикулы содержат нейропептиды. Агранулярные пузырьки содержат ацетилхолин. Медиаторами возбуждения могут быть также производные глутаминовой и аспарагиновой кислот. Действие медиатора на постсинаптическую мембрану заключается в повышении ее проницаемости для ионов Na+. Возникновение потока ионов Na+ из синаптической щели через постсинаптическую мембрану ведет к ее деполяризации и вызывает генерацию возбуждающего постсинаптического потенциала (ВПСП). Для синапсов с химическим способом передачи возбуждения характерны синоптическая задержка проведения возбуждения, длящаяся около 0,5 мс, и развитие постсинаптического потенциала в ответ на пресинаптический импульс. Этот потенциал при возбуждении проявляется в деполяризации постсинаптической мембраны (ВПСП), а при торможении — в гиперполяризации ее, в результате чего развивается тормозной постсинаптический потенциал (ТПСП). При возбуждении проводимость постсинаптической мембраны увеличивается. ВПСП возникает в нейронах при действии в синапсах ацетилхолина, норадреналина, дофамина, серотонина, глутаминовой кислоты, вещества Р. Величина ВПСП зависит от количества выделившегося медиатора и может составлять 0,12—5,0 мВ. Под влиянием ВПСП деполяризуются соседние с синапсом участки мембраны, затем деполяризация достигает аксонного холмика нейрона, где возникает возбуждение, распространяющееся на аксон. ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. Они изменяют ионную проницаемость постсинаптической мембраны таким образом, что в ней открываются поры диаметром около 0,5 нм. Эти поры не пропускают ионы Na+ (что вызвало бы деполяризацию мембраны), но пропускают ионы К+ из клетки наружу, в результате чего происходит гиперполяризация постсинаптической мембраны. Переход медиатора в синаптическую щель осуществляется путем экзоцитоза: пузырек с медиатором соприкасается и сливается с пресинаптической мембраной, затем открывается выход в синаптическую щель и в нее попадает медиатор. В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего ПД количество медиатора резко возрастает. Затем медиатор перемещается к постсинаптической мембране, действует на специфические для него рецепторы и образует на мембране комплекс медиатор—рецептор. Данный комплекс изменяет проницаемость мембраны для ионов К+ и Na+, в результате чего изменяется ее потенциал покоя. В пресинаптической терминали образуется и скапливается в виде пузырьков медиатор, например, ацетилхолин. В каждом пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант нейромедиатора. При возбуждении пресинаптической части синапса идущим по аксону электрическим импульсом, ее мембрана становится проницаемой для ацетилхолина. Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. В пресинаптическую мембрану встроены потенциалозависимые Ca2+ каналы. При поступлении ПД к терминальному расширению мембрана деполяризуется, Ca2+ каналы открываются, ионы Ca2+ из синаптической щели входят в терминаль, запуская в активных зонах процесс слияния мембраны синаптического пузырька и пресинаптической мембраны, т.е. секрецию (экзоцитоз) нейромедиатора. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе путём реорганизации примембранного цитоскелета). Ацетилхолин высвобождается и проникает в синаптическую щель. Здесь он взаимодействует со своими рецепторами постсинаптической мембраны, принадлежащей мышечному волокну. Рецепторы, возбуждаясь, открывают белковый канал, встроенный в липидный слой мембраны. Через открытый канал внутрь мышечной клетки проникают ионы Na+, что приводит к деполяризации мембраны и возникновению ВПСП. Он вызывает генерацию потенциала действия в соседних электрогенных участках мембраны нейрона Этапы синаптической передачи. Синаптическая передача возможна при реализации ряда последовательных процессов: синтез нейромедиатора, его накопление и хранение в синаптических пузырьках вблизи пресинаптической мембраны, высвобождение нейромедиатора из нервной терминали, кратковременное взаимодействие нейромедиатора с рецептором, встроенным в постсинаптическую мембрану; разрушение нейромедиатора или захват его нервной терминалью. На рис. 11 представлены стандартные этапы синаптической передачи. Р 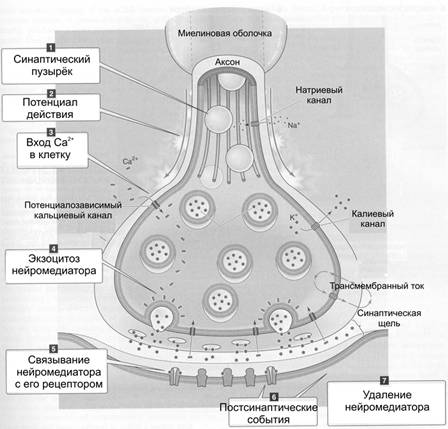 ис. 11. Этапы синаптической передачи ис. 11. Этапы синаптической передачи1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и концентрирующиеся в активных зонах пресинаптической мембраны. 2. Приходящий по аксону ПД деполяризует пресинаптическую мембрану. 3. Вследствие деполяризации открываются потенциалозависимые Са2+ каналы, и Са2+ поступает в терминаль. 4. Увеличение внутриклеточного Са2+ запускает слияние синаптических пузырьков с пресинаптической мембраной и выброс нейромедиатора в синаптическую щель (экзоцитоз). 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными для них рецепторами постсинаптической мембраны. 6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов через ионотропные рецепторы) либо опосредованно — активация ионных каналов через систему G белка (метаботропные рецепторы). 7. Инактивация нейромедиаторов происходит либо путём их ферментной деградации, либо молекулы нейромедиатора захватываются клетками. Таким образом, синаптическая передача — сложный каскад событий. Многие неврологические и психиатрические заболевания сопровождаются нарушением синаптической передачи. Различные ЛС влияют на синаптическую передачу, вызывая нежелательный эффект (например, галлюциногены) или, наоборот, корригируя патологический процесс (например, психофармаколо-гические антипсихотические средства). Характеристика отдельных нейромедиаторов Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон, постганглионарных холинергических (парасимпатических) волокон вегетативной нервной системы и разветвлений аксонов многих нейронов ЦНС (базальные ганглии, двигательная кора). Синтезируется из холина и ацетил-КоА при помощи холинацетилтрансферазы, взаимодействует с холинорецепторами нескольких типов. Кратковременное взаимодействие лиганда с рецептором прекращает ацетилхолинэстераза, гидролизующая ацетилхолин на холин и ацетат. Дофамин — нейромедиатор в окончаниях некоторых аксонов периферических нервов и многих нейронов ЦНС (чёрное вещество, средний мозг, гипоталамус). После секреции и взаимодействия с рецепторами дофамин активно захватывается пресинаптической терминалью, где его расщепляет моноаминооксидаза. Дофамин метаболизирует с образованием ряда веществ, в т.ч. гомованилиновой кислоты. Норадреналин секретируется из большинства постганглионарных симпатических волокон и является нейромедиатором между многими нейронами ЦНС (например, гипоталамус, голубоватое место). Образуется из дофамина путём гидролиза при помощи дофамин-b-гидроксилазы. Норадреналин хранится в синаптических пузырьках, после высвобождения взаимодействует с адренорецепторами, реакция прекращается в результате захвата норадреналина пресинаптической частью. Уровень норадреналина определяется активностью тирозин гидроксилазы и моноаминооксидазы. Норадреналин — мощный вазоконстриктор, эффект происходит при взаимодействии нейромедиатора с ГМК стенки кровеносных сосудов. Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва, нейроны восходящей ретикулярной активирующей системы). Предшественником служит триптофан, гидроксилируемый триптофангидроксилазой до 5-гидрокситриптофана с последующим декарбоксилированием декарбоксилазой L-аминокислот. Расщепляется моноаминооксидазой с образованием 5-гидроксииндолуксусной кислоты. g-Аминомасляная кислота — тормозный нейромедиатор в ЦНС (базальные ганглии, мозжечок). Образуется из глутаминовой кислоты под действием декарбоксилазы глутаминовой кислоты, захватывается из межклеточного пространства пресинаптической частью и деградирует под влиянием трансаминазы ГАМК. b-Эндорфин — нейромедиатор полипептидной природы многих нейронов ЦНС (гипоталамус, миндалина мозжечка, таламус, голубоватое место). Метионин–энкефалин и лейцин–энкефалин — небольшие пептиды (5 аминокислотных остатков), присутствующие во многих нейронах ЦНС (бледный шар, таламус, хвостатое ядро, центральное серое вещество). Как и эндорфин, образуются из проопиомеланокортина. После секреции взаимодействуют с пептидергическими (опиоидными) рецепторами. Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые присутствуют в нейронах тех же анатомических областей, что и энкефалинергические нейроны. Образуются из продинорфина, инактивируются путём гидролиза. Вещество Р — нейромедиатор пептидной природы в нейронах центральной и периферической нервной системы (базальные ганглии, гипоталамус, спинномозговые узлы). Передача болевых стимулов реализуется при помощи вещества Р и опиоидных пептидов. Глицин, глутаминовая и аспарагиновая кислоты. Эти аминокислоты в некоторых синапсах являются нейромедиаторами (глицин во вставочных нейронах спинного мозга, глутаминовая кислота — в нейронах мозжечка и спинного мозга, аспарагиновая кислота — в нейронах коры). Глутаминовая и аспарагиновая кислоты вызывают возбуждающие ответы, а глицин — тормозные. Свойства синапсов в ЦНС 1) Относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий медиатор; 2) наличие хемочувствительных рецепторуправляемых каналов постсинаптической мембраны; 3) возможность действия специфических блокирующих агентов на рецептирующие структуры постсинаптической мембраны; 4) увеличение длительности постсинаптического потенциала мембраны при подавлении действия ферментов, разрушающих синаптической медиатор; 5) развитие в постсинаптической мембране ПСП из миниатюрных потенциалов, обусловленных квантами медиатора; 6) зависимость длительности активной фазы действия медиатора в синапсе от свойств медиатора; 7) односторонность проведения возбуждения; 8) увеличение выделения квантов медиатора в синаптическую щель пропорционально частоте приходящих по аксону импульсов; 9) зависимость увеличения эффективности синаптической передачи от частоты использования синапса («эффект тренировки»); 10) утомляемость синапса, развивающаяся в результате длительного высокочастотного его стимулирования. В этом случае утомление может быть обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией постсинаптической мембраны (пессимальное торможение). Перечисленные свойства относятся к химическим синапсам. Электрические синапсы имеют некоторые особенности, а именно: малую задержку проведения возбуждения; возникновение деполяризации как в пре-, так и в постсинаптической частях синапса; наличие большей площади синаптической щели в электрическом синапсе, чем в химическом. Электрические синапсы. Помимо химической передачи в синапсах, между нейронами, а также нейронами и глиоцитами возможна и электрическая передача возбуждения — при помощи щелевых контактов, обеспечивающих электротоническое, метаболическое и информационное сообщение между контактирующими клетками. По отношению к таким межклеточным контактам между нервными элементами применяют термин «эфапс» (греч. ephapsis — прикосновение, касание, контакт). Такие электрические синапсы широко распространены в нервной системе беспозвоночных, но изредка встречаются и в ЦНС млекопитающих. Так, в сетчатке глаза горизонтальные клетки связаны между собой щелевыми контактами. Благодаря этому локальные потенциалы, потенциалы действия, а также продукты метаболизма и красители (в эксперименте) могут распространяться от клетки к клетке, оказывая значительное влияние на процесс переработки зрительной информации. Основные отличия между химическими и электрическими синапсами представлены в таблице. Химические и электрические контакты между нейронами
|