Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

4-2. Современная теория мышечного сокращения и расслабления. Электромеханическое сопряжение.Механизм мышечного сокращения. В присутствии АТФ, но при низкой концентрации Са2+, составляющей 10–8 моль/л и меньше, волокно находится в расслабленном состоянии, т.е. головки миозина не образуют связи с актином. В момент увеличения концентрации Са2+ вблизи головок до 10–6-10–5 моль/л ферментативная активность головок многократно увеличивается и в той же мере возрастает гидролиз АТФ. В скелетных мышцах тяжи тропомиозина закрывают активные участки актинового филамента, препятствуя взаимодействию миозиновых головок с мономерами актина, тем самым предотвращая сокращение. Повышение концентрации Са2+ сопровождается его связыванием с тропонином. При этом молекулы комплекса тропонин-тропомиозин изменяют свое расположение таким образом, что обнажают активные участки на актиновом филаменте. За счет освобождающейся энергии происходит связывание головок с нитью актина, изгибание «ножки» и вследствие этого актиновые филаменты продольно перемещаются относительно центрального миозинового стержня. Уменьшение же концентрации Са2+ до прежнего минимального уровня вызывает размыкание поперечных мостиков и возвращение Описанное взаимодействие глобулярной головки миозина и активным участком молекулы актина называют циклом поперечного мостика. Один цикл поперечного мостика происходит в следующей последовательности: активация миозиновой головки; обнажение активного участка молекулы актина в присутствии кальция; самопроизвольное формирование поперечного мостика; поворот глобулярной головки, сопровождающийся продвижением актиновой нити и укорочением саркомера; разрыв поперечного мостика. Цикл после завершения может повториться или остановиться. Если бы все поперечные мостики в отдельной мышце прошли одновременно только один цикл, мышца укоротилась бы всего на 1% от ее длины в состоянии покоя. Однако многие мышцы способны укорачиваться до 60% длины покоя. Это показывает, что цикл поперечного мостика происходит в них многократно. При этом каждый раз миозин захватывает и тянет новый участок актина. Именно такая многократность повторения цикла замыкания-размыкания мостика вызывает более выраженное укорочение и генерирует бόльшую силу. 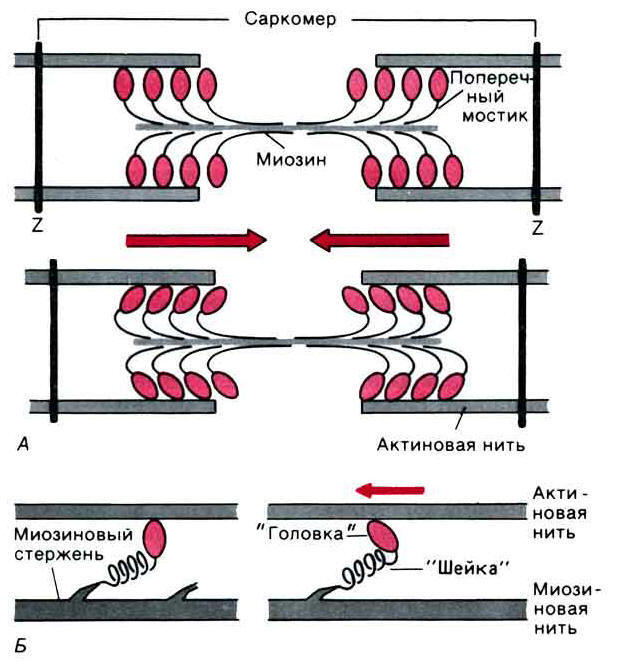 Рис. 7. Функция поперечных мостиков. А. Модель механизма сокращения: миозиновая нить с поперечными мостиками, прикрепленными к соседним актиновым нитям; вверху - до, внизу - после "гребка" мостиков (на самом деле они функционируют асинхронно). Б. Модель генерирования силы поперечными мостиками; слева - до, справа - после "гребка". Поперечные мостики химически соответствуют субфрагменту миозина - "тяжелому меромиозину", который состоит из субфрагментов I (головка) и II (ножка). В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются к ее центру. Прочная соединительнотканная фасциальная оболочка, прикрепляющая мышцу посредством сухожилия к костям, передает вызываемую сокращением силу костям. Линейно направленная мышечная сила тянет кость и вызывает ее вращение в плоскости, перпендикулярной оси сустава. Поскольку мышца обычно проходит, по крайней мере, через один сустав, более свободный сегмент костного скелета будет перемещаться, вызывая вращательное движение в суставе. Электромеханическое сопряжение. Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала о начале сокращения от возбужденной сарколеммы, называется электромеханическим сопряжением. Электромеханическое сопряжение происходит посредством распространения возбуждения (потенциала действия) по системе поперечных трубочек внутрь волокна. При этом информация о возбуждении (фактически о перезарядке мембраны) очень быстро достигает продольной системы СПР, в конечном итоге вызывая открытие в мембране цистерн Са2+ -каналов и залповый выброс катиона, быстро диффундирующего к миофибриллам. Ключевое значение в процессе передачи этого сигнала имеет разница электрических потенциалов по разные стороны мембраны Т-трубочки. Предполагают, что развивающаяся при передаче потенциала действия по Т-трубочке деполяризация вызывает конформационные изменения белков ножки концевой цистерны. Эти конформационные изменения приводят к открытию Са2+ -каналов в СПР и обеспечивают выход ионов Са2+ в цитоплазму по электрохимическому градиенту. Остановка сокращения. Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного импульса в двигательной концевой пластинке нервно-мышечного соединения. Прекращение импульса означает устранение сигнала к высвобождению Са2+, которое прекращается. Однако действие остающегося в саркоплазме Са2+ может все еще продолжаться. Поэтому при одиночном нервном импульсе и соответственно одиночном сокращении генерация силы (укорочение) саркомером только останавливается. Чтобы сила, генерируемая саркомером, уменьшилась до величины в состоянии покоя, и произошло расслабление саркомера, необходимо многократное уменьшение Са2+ вблизи миозиновых головок. Это происходит вследствие откачки Са2+, усиливающейся по мере его выхода в саркоплазму, поскольку катионы Са2+ все сильнее начинают активировать Са2+ -насос (Са2+ -АТФазу) мембран саркоплазматического ретикулума. В результате «накачивания» Са2+ в цистерны СПР его концентрация вблизи головок резко уменьшается и поперечные мостики размыкаются – мышечное волокно расслабляется. Циклическое образование поперечных мостиков продолжается до тех пор, пока в волокне имеются в достаточном количестве свободных катионов кальция и АТФ. Все, что снижает концентрацию Са2+ и АТФ, будет ослаблять мышечное сокращение, что происходит при утомлении мышцы. Таковыми факторами, например, является низкая концентрация калия, яды и токсины, нарушающие выработку АТФ в окислительном фосфорилировании. Полное отсутствие АТФ вызывает устойчивое сокращение, называемое трупным окоченением. Такое состояния устойчивого сокращения обусловлено двумя обстоятельствами. Во-первых, в отсутствие АТФ невозможна откачка Са2+ и саркоплазмы в везикулы СПР. Во-вторых, без АТФ не может произойти повторная активация миозиновой головки, требующаяся для разрыва поперечного мостика между актином и миозином. В итоге поперечные мостики остаются замкнутыми, а мышца в сокращенном состоянии. |