Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

3.2. Нервно-мышечный синапс: строение, механизм проведения возбуждения, особенности проведения возбуждения в синапсе по сравнению с нервным волокном.Синапсами называются контакты, которые устанавливают нейроны как самостоятельные образования. Синапс представляет собой сложную структуру и состоит из пресинаптической части (окончание аксона, передающее сигнал), синаптической щели и постсинаптической части (структура воспринимающей клетки). Р 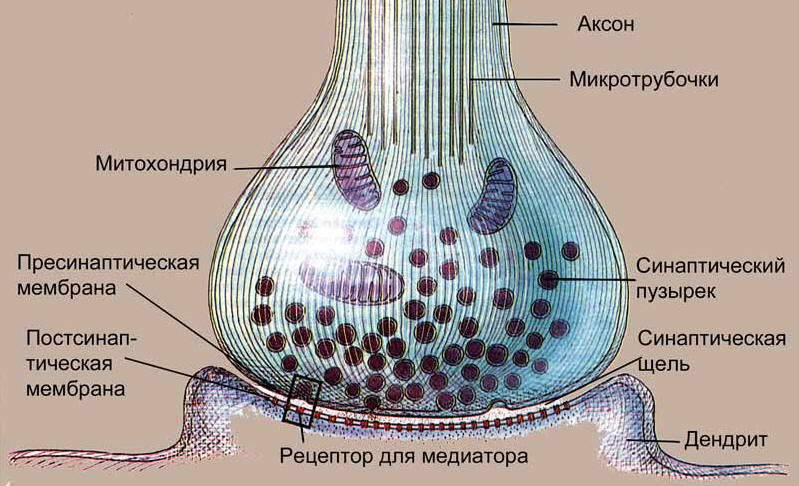 ис. 5. Схема строения синапса ис. 5. Схема строения синапсаПресинаптическая часть химического синапса образуется расширением аксона по его ходу или окончания. В пресинаптической части имеются пузырьки. Пузырьки (кванты) содержат медиатор. В пресинаптическом расширении находятся митохондрии, обеспечивающие синтез медиатора, гранулы гликогена и др. При многократном раздражении пресинаптического окончания запасы медиатора в синаптических пузырьках истощаются. Пузырьки содержат ацетилхолин, который является медиатором в нерно-мышечных синапсах (рис. 5). Роль медиатора заключается в повышении проницаемости постсинаптической мембраны для ионов Na+. Возникновение потока ионов Na+ из синаптической щели через постсинаптическую мембрану ведет к ее деполяризации и вызывает генерацию возбуждающего постсинап-тического потенциала (ВПСП). Для распространения возбуждения через химический синапс важно, что нервный импульс, идущий по пресинаптической части, полностью гасится в синаптической щели. Однако этот импульс вызывает физиологические изменения в пресинаптической части мембраны. В результате у ее поверхности скапливаются синаптические пузырьки, изливающие медиатор в синаптическую щель. Переход медиатора в синаптическую щель осуществляется путем экзоцитоза: пузырек с медиатором соприкасается и сливается с пресинаптической мембраной, затем открывается выход в синаптическую щель и в нее попадает медиатор. В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего возбуждения количество медиатора резко возрастает. Нервно-мышечные синапсы обеспечивают проведение возбуждения с нервного волокна на мышечное благодаря медиатору ацетилхолину, который при возбуждении нервного окончания переходит в синаптическую щель и действует на концевую пластинку мышечного волокна. В пресинаптической терминали образуется и скапливается в виде пузырьков ацетилхолин. При возбуждении электрическим импульсом, идущим по аксону, пресинаптической части синапса ее мембрана становится проницаемой для ацетилхолина. Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. Ион Са2+ входит в пресинаптическую часть синапса из синаптической щели. Ацетилхолин высвобождается и проникает в синаптическую щель. Здесь он взаимодействует со своими рецепторами постсинаптической мембраны, принадлежащей мышечному волокну. Рецепторы, возбуждаясь, открывают белковый канал, встроенный в липидный слой мембраны. Через открытый канал внутрь мышечной клетки проникают ионы Na+, что приводит к деполяризации мембраны мышечной клетки, в результате развивается так называемый потенциал концевой пластинки (ПКП). Он вызывает генерацию потенциала действия мышечного волокна. Нервно-мышечный синапс передает возбуждение в одном направлении: от нервного окончания к постсинаптической мембране мышечного волокна, что обусловлено наличием химического звена в механизме нервно-мышечной передачи. Скорость проведения возбуждения через синапс намного меньше, чем по нервному волокну, так как здесь тратится время на активацию пресинаптической мембраны, переход через нее кальция, выделение ацетилхолина в синаптическую щель, деполяризацию постсинаптической мембраны, развитие ПКП. Синаптическая передача возбуждения по сравнению с распространением потенциала действия имеет рад свойств: 1) наличие медиатора в пресинаптической части синапса; 2) относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий медиатор; 3) возможность действия специфических блокирующих агентов на рецептирующие структуры постсинаптической мембраны;\ 4) зависимость длительности активной фазы действия медиатора в синапсе от свойств медиатора; 5) односторонность проведения возбуждения; 6) наличие хемочувствительных рецепторуправляемых каналов постсинаптической мембраны; 7) увеличение выделения квантов медиатора в синаптическую щель пропорционально частоте приходящих по аксону импульсов; 8) зависимость увеличения эффективности синаптической передачи от частоты использования синапса («эффект тренировки»; 9) утомляемость синапса, развивающаяся в результате длительного высокочастотного его стимулирования. В этом случае утомление может быть обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса. Синаптические медиаторы являются веществами, которые имеют специфические инактиваторы. Например, ацетилхолин инактивируется ацетилхолинэстеразой. Неиспользованный медиатор и его фрагменты всасываются обратно в пресинаптическую часть синапса. В нервно-мышечном синапсе в норме ацетилхолин действует на синаптическую мембрану короткое время (1—2 мс), так как сразу же начинает разрушаться ацетилхолинэстеразой. В случаях, когда этого не происходит и ацетилхолин не разрушается на протяжении сотни миллисекунд, его действие на мембрану прекращается и мембрана не деполяризуется, а гиперполяризуется и возбуждение через этот синапс блокируется. Специально для снижения тонуса мышц, особенно при операциях, используют блокаду нервно-мышечной передачи, которая может быть вызвана следующими способами: 1) действие местноанестезирующих веществ, которые блокируют возбуждение в пресинаптической части; 2) блокада высвобождения медиатора в пресинаптической части (например, ботулинический токсин); 3) нарушение синтеза медиатора, например при действии гемихолиния; 4) блокада рецепторов ацетилхолина, например при действии бунгаротоксина; 5) вытеснение ацетилхолина из рецепторов, например действие кураре; 6) инактивация постсинаптической мембраны сукцинилхолином и др.; 7) угнетение холинэстеразы, что приводит к длительному сохранению ацетилхолина и вызывает глубокую деполяризацию и инактивацию рецепторов синапсов. Такой эффект наблюдается при действии фосфорорганических соединений. |