Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

2.2. Потенциал действия: фазы потенциала действия, механизм возникновения. Восстановительный период. Явление аккомодации возбудимой ткани.Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя (например, толчка электрического тока), в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД) Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+ превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно. При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+. Поэтому поток ионов Na+ в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+ достигает величины +150 мв. Одновременно несколько уменьшается выход К+ из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации). При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. "перезарядка мембраны". Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне. Потенциал действия протекает фазно. Временной ход потенциала действия включает четыре последовательных этапа: локальный ответ, деполяризацию, реполяризацию и следовые потенциалы (рис. 2). В ПД принято различать его пик (т.н. спайк - spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее. 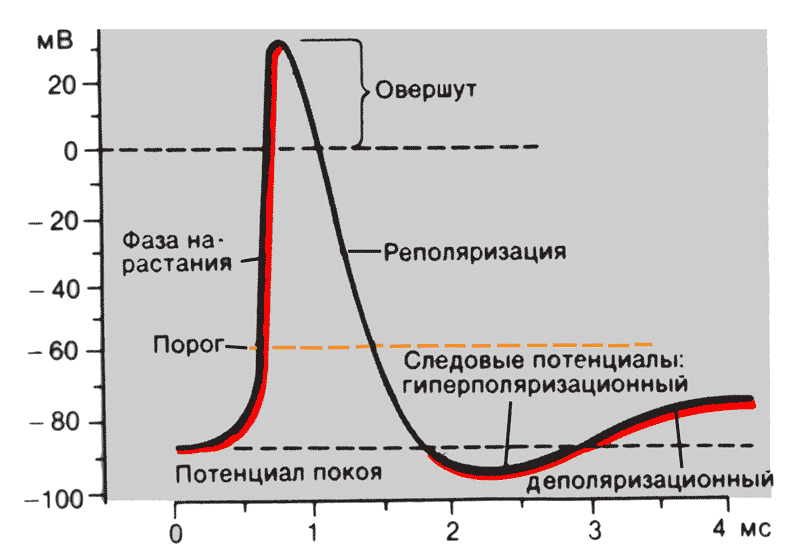 Рис. 2. Фазы и временной ход потенциала действия. Кроме пика, в ПД различают два следовых потенциала - следовую деполяризацию (следовой отрица-тельный потенциал) и следовую гиперполяризацию (следовой положи-тельный потенциал. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения. Промежуток времени, в течение которого сохраняется активное состояние в виде ПД, неодинаков в разных возбудимых структурах. В нейронах он составляет около 1 мс, в волокнах скелетных мышц – 10 мс, в миокарде достигает 200–250 мс. Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону называется деполяризацией. Область электроположительности носит название овершута, правое крыло ПД, свидетельствующее о восстановлении исходного поляризованного состояния мембраны принято называть реполяризацией. Часто, но не всегда возвращение ПД к исходному уровню в состоянии покоя происходит с наличием фаз в форме так называемых следовых потенциалов. Следовые потенциалы неодинаковы в мышцах и нервах. В волокнах скелетных мышц фаза реполяризации очень замедлена. Примерно через 1 мс после начала ПД наблюдается отчетливый перегиб крыла реполяризации – это следовая деполяризация. В нейроне чаще всего кривая реполяризации быстро пересекает уровень МПП и на некоторое время потенциал мембраны становится более электроотрицательным, чем МП. Это явление называют следовой гиперполяризацией. Повышение проницаемости мембраны для ионов Na+ продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается. Наличие специальных Na- и К- каналов и сложного механизма запирания и открытия ворот изучено биофизиками достаточно хорошо. Показано, что существуют избирательные механизмы, регулирующие те или иные каналы. Например, яд тетродотоксин блокирует только Na-поры, а тетраэтиламмоний - только К-поры. Показано, что у некоторых клеток возникновение возбуждения связано в изменением проницаемости мембраны для Са++, в других - для Mg+. Исследования механизмов изменения проницаемости мембран продолжаются. В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса. В результате повышения проводимости резко возрастает поток катионов Na+, поэтому отрицательный заряд в клетке вблизи внутренней стороны поверхности мембраны также резко уменьшается вплоть до преобладания положительных зарядов. В результате происходит изменение знака потенциала, достигающего +30 мВ. После этого проводимость мембраны для Na+ также резко снижается. Для нормального протекания ПД играет существенную роль и изменение проводимости мембраны для K+, которая начинает возрастать позже возрастания проводимости для Na+. Увеличение относительно медленного выхода K+ из клетки в фазу снижения проводимости для Na+ вызывает реполяризацию мембраны. Таким образом, в живой клетке существуют два различных типа движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным транспортом. Он ответственен за возникновение МП и ПД и ведет в конечном итоге к выравниванию концентраций ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в "выкачивании" ионов Na+ из протоплазмы и "нагнетании" ионов К+ внутрь клетки. Этот тип ионного транспорта возможет лишь при условии затраты энергии - это активный транспорт. Он является результатом работы специальных ферментных систем (т.н. насосов), и благодаря ему восстанавливается исходная разность концентраций, необходимая для поддержания МП. Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше - надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 - 10-9 А. Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк) Инактивация Na+-системы. Na+-системой обозначают механизм, позволяющий в течение нескольких долей миллисекунды многократно (до 20 раз) увеличить проводимость клеточной мембраны для Na+. Достигнув пикового значения, примерно через 0,5 мс проводимость мембраны для Na+ начинает снижаться. Быстрое снижение проводимости для Na+ называют инактивацией Na+-системы. В основе инактивации Na+-системы лежит переход в инактивационное состояние потенциалзависимых Na+-каналов. Поэтому скорость и степень снижения проводимости потенциалзависимы. Это означает, что чем больше отличается потенциал мембраны от мембранного потенциала покоя в сторону электроположительности, тем сильнее инактивирована Na+-система. Поэтому деполяризация мембраны вызывает снижение тока Na+ внутрь клетки. С одной стороны, это свидетельствует о том, что усиление тока Na+ само себе служит причиной его быстрого последующего снижения и начала развития реполяризации. С другой стороны, это означает, что если исходный потенциал клетки выше потенциала покоя на 20–30 мВ, то Na+-система полностью инактивирована и никакая последующая деполяризация уже не может активировать ее, т.е. вызвать резкое увеличение проводимости для Na+ и генерацию ПД. |