Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

2. 3. Параметры возбудимости ткани: порог, полезное время и хронаксия, критический наклон, лабильность.Зависимость пороговой силы стимула от его длительности. Как уже указывалось, пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. В особенно четкой форме эта зависимость проявляется при использовании в качестве раздражителя прямоугольных толчков постоянного тока. Полученная в таких опытах кривая получила название "кривой силы-времени" (рис. 3). Она была изучена Гоорвегом, Вейсом и Лапиком в начале века. Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной величины или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. При очень коротких стимулах кривая силы-времени становится параллельной оси координат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражения.  Рис. 3. Кривая "силы-времени Гоорвега - Вейса Определение полезного времени практически затруднено, так как точка полезного времени находится на участке кривой, переходящей в параллельную. Поэтому Лапик предложил использовать полезное время двух реобаз - хронаксию. Ее точка находится на самом крутом участке кривой Гоорвега-Вейса. Хронаксиметрия получила широкое распространение как в эксперименте, так и в клинике для диагностики повреждений волокон двигательных нервов. Таким образом, второй параметр возбудимости тканей - полезное время (хронаксия). Зависимость порога от крутизны нарастания силы раздражителя. Величина порога раздражения нерва или мышцы зависит не только от длительности стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием тока. Если же вместо таких стимулов применять линейно или экспоненциально нарастающие стимулы, пороги оказываются увеличенными и тем больше, чем медленнее нарастает ток. При уменьшении крутизны нарастания тока ниже некоторой минимальной величины (т.н. критический наклон) ПД вообще не возникает, до какой бы конечной силы не увеличивался ток. Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю получило название аккомодация. Чем выше скорость аккомодации, тем более круто должен нарастать стимул, чтобы не утратить своего раздражающего действия. Порог тем выше, чем меньше критический наклон. Аккомодация к медленно нарастающему току обусловлена тем, что за время действия этого тока в мембране успевают развиться процессы, препятствующие возникновению ПД. Дело в том, что деполяризация мембраны одновременно приводит к началу двух процессов: одного - быстрого, ведущего к повышению натриевой проницаемости и возникновению ПД, а другого - медленного, приводящего к инактивации натриевой проницаемости и к окончанию возбуждения. При крутом нарастании стимула Na-активация успевает достичь значительной величины прежде, чем развивается Na-инактивация. В случае медленного нарастания силы тока на первый план выступают процессы инактивации, приводящие к повышению порога и снижению амплитуды ПД. Все агенты, усиливающие или ускоряющие инактивацию, увеличивают скорость аккомодации. В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой стимул еще сохраняет способность вызывать ПД. Эту минимальную крутизну называют критическим наклоном. Его выражают или в абсолютных единицах (мА/сек), или в относительных (как отношение пороговой силы того постепенно нарастающего тока, который еще способен вызывать возбуждение, к реобазе прямоугольного толчка тока). Таким образом, третий параметр возбудимости - критический наклон. Четвертый параметр возбудимости - лабильность (функциональная подвижность). Под функциональной подвижностью понимают скорость протекания элементарных физиологических реакций, с которой система успевает перейти от состояния возбуждения к состоянию покоя. Иначе говоря, чем большее число раз в единицу времени система способна прийти в состояние возбуждения, тем ее функциональная подвижность выше. Поскольку каждая система обладает своей длительностью возбуждения, т.е. интервалом времени, в течение которого оно существует, то подвижность разных структур неодинакова. Лабильность измеряется максимальным числом импульсов, которое ткань может воспроизвести в соответствии с частотой раздражения и зависит от длительности рефрактерности. Сравнительная характеристика возбудимости разных тканей. Самой высокой возбудимостью по всем параметрам обладает нервная ткань, на втором месте - скелетная мускулатура, на третьем месте - гладкая мускулатура, и, наконец, наименее всего возбудимы секреторные клетки. 2-4. Сравнительная характеристика локального потенциала и потенциала действия. Закон "Все или ничего". Фазовые изменения возбудимости нерва во время возбуждения (потенциала действия), их механизм. Возникновение и развитие местного потенциала отличается определенными и специфическими особенностями: 1. Местный потенциал возникает при действии подпороговых раздражителей малой силы. 2. Амплитуда местного потенциала градуально зависит от силы раздражителя – чем сильнее раздражение, тем больше амплитуда потенциала. 3. Продолжительность местного потенциала непостоянна и зависит от силы и длительности раздражителя. Чем сильнее и продолжительнее раздражение, тем дольше сохраняется местный потенциал. 4. У местного потенциала отсутствует латентный период – он возникает практически сразу после воздействия раздражителя. 5. Участок мембраны, в границах которого возникает местный потенциал, переходит в состояние повышенной возбудимости. 6. Местные потенциалы могут суммироваться. Если на фоне местного потенциала нанести новое подпороговое раздражение, возникший на второе раздражение ответ накладывается на первый, и общий суммарный эффект от этого увеличивается Закон "все или ничего". При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н. закон "все или ничего". Согласно этому закону, под пороговые раздражения не вызывают возбуждения ("ничего"), при пороговых же стимулах возбуждение сразу приобретает максимальную величину ("все"), и уже не возрастает при дальнейшем усилении раздражителя. Долгое время закон "все или ничего" неправильно интерпретировался как общий принцип реагирования возбудимых тканей. Предполагали, что "ничего" означает полное отсутствие ответа на под пороговый стимул, а "все" рассматривалось как проявление полного исчерпания возбудимым субстратом его потенциальных возможностей. Дальнейшие исследования, в особенности микроэлектродные, показали, что эта точка зрения не соответствует действительности. Выяснилось, что при под пороговых силах возникает местное не распространяющееся возбуждение (локальный ответ). Вместе с тем оказалось, что "все" также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу "все или ничего" - его амплитуда начинает градуально зависеть от силы стимула. Поэтому "все или ничего" рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях. Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для их изучения нерв или мышцу подвергают действию двух коротких электрических стимулов, следующих друг за другом с определенным интервалом. Первый называется раздражающим, второй - тестирующим. Регистрация возникающих в ответ на эти раздражения ПД позволила установить важные факты 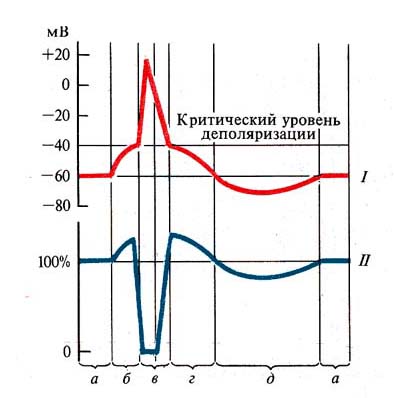 Рис. 4. Сопоставление одиночного возбуждения (/) с фазами возбудимости (//): а — мембранный потенциал (исходная возбудимость), б — локальный ответ, или ВПСП (повышенная возбудимость), в — потенциал действия (абсолютная и относительная рефрактерность), г — следовая деполяризация (супернормальная возбудимость), д — следовая гиперполяризация (субнормальная возбудимость) Во время локального ответа возбудимость повышена, так как мембрана деполяризована и разность между Е0 и Ек падает. Периоду же возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название абсолютной рефрактерности (невпечатлительности). В это время тестирующий стимул не способен вызвать новый ПД, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности примерно совпадает с длительностью восходящей ветви ПД. В быстро проводящих нервных волокнах она составляет 0,4-0,7 мсек. В волокнах мышцы сердца - 250-300 мсек. Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена. Согласно ионной теории Ходжкина-Хаксли, абсолютная рефрактерность обусловлена вначале наличием максимальной натриевой проницаемости, когда новый стимул не может что-то изменить или добавить, а затем развитием натриевой инактивации, закрывающей Na-каналы. Вслед за этим происходит снижение натриевой инактивации, в результате чего постепенно восстанавливается способность волокна генерировать ПД. Это - состояние относительной рефрактерности. Относительная рефрактерная фаза сменяется фазой повышенной (супернормальной) возбудимости, совпадающей по времени с периодом следовой деполяризации. В это время разность между Ео и Ек ниже исходной. В двигательных нервных волокнах теплокровных животных длительность супернормальной фазы составляет 12-30 мсек. Период повышенной возбудимости сменяется субнормальной фазой , которая совпадает со следовой гиперполяризацией. В это время разница между мембранным потенциалом (Ео) и критическим уровнем деполяризации (Ек) увеличивается. Длительность этой фазы составляет несколько десятков или сотен мсек. |