Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

Лекция 4. ФИЗИОЛОГИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ4-1. Скелетная мышца: строение мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы, физиологические свойства скелетной мышцы и ее функции.Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц – мышечных волокон или мышечных клеток. Они имеют цилиндрическую форму и расположены параллельно друг другу. Волокна состоят из большого количества миофибрилл, создающих характерную поперечно-полосатую исчерченность. Миофибрилла включает белковые нити двух типов – актиновые (тонкие) и миозиновые(толстые). Между миофибриллами находится множество митохондрий. Цитоплазма волокна или саркоплазма содержит сеть внутренних полостей, окруженных мембранами – саркоплазматический ретикулум (СПР). Отдельная замкнутая везикула СПР часто называется цистерной. Поперек волокна между миофибриллами проходит система трубочек, представляющих впячивания сарколеммы, называемые Т-системой. В определенных местах трубочки располагаются между двумя цистернами. Комплекс, состоящий из одной Т-трубочки и двух, тесно примыкающих к ней цистерн, представляет триаду. Трубочки и цистерны соединены между собой поперечными мембранными мостиками. Саркоплазматический ретикулум, являясь хранилищем Са2+, выполняет функцию регулятора уровня свободных катионов Са2+ в саркоплазме. В световой микроскоп видна только поперечная исчерченность миофибрилл, включающая светлый диск I, темный диск А и Z–линию, разделяющую зону I. Участок между двумя полосками Z называется саркомером. При гораздо большем увеличении, которое дает электронный микроскоп, видно, что такая картина полосатости обусловлена определенным расположением нитей актина и миозина. Молекула миозина состоит из длинного участка – хвоста и глобулярной части из двух головок. Молекулы миозина так расположены в миозиновой нити, что головки равномерно распределены по ее длине, кроме небольшого участка в середине, где они отсутствуют. Для механизма сокращения принципиально, что в местах перекрытия нитей актина и головок миозина они могут взаимодействовать между собой, вызывая укорочение мышцы. Энергия для совершения такой работы черпается из АТФ, поэтому неудивительно, что миозиновые головки обладают АТФ-азной активностью. 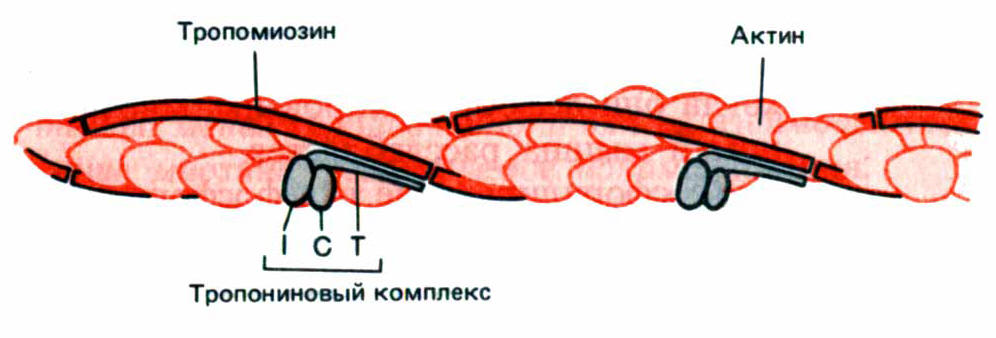 Рис. 6. Схематическое изображение тонкой актиновой нити , показывающее локализацию тропомиозина и тропонина на его поверхности. I, C, T - белок тропонин, состоящий из трех полипептидных цепей (тропонины I, C, T) Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет участок для связывания тропомиозина и ответственен за прикрепление всего комплекса к актиновому филаменту. Если к тропонину Т и тропомиозину добавить тропонин I, образуется комплекс, препятствующий взаимодействию актина с миозином даже в присутстви катионов Са2+. Дополненный тропонином С, этот комплекс продолжает блокировать актин-миозиновое взаимодействие, но только в отсутствии Са2+ ; при наличии в среде свободных катионов кальция тропонин С связывает четыре катиона Са2+ и таком состоянии снимает блокаду. именно этот эффект и лежит в основе индукции мышечного сокращения катионами кальция. Актиновые филаменты имеют сложный состав и структуру. Каждая актиновая нить состоит из двух спиральных тяжей, каждый из которых образован молекулами глобулярного G-актина. Комплекс из двух нитей называется F-актином. Кроме того, в актиновом филаменте содержатся вспомогательные регуляторные белки тропомиозин и тропонин. Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные поперечно-полосатые мышцы подразделяют на тонические и фазные волокна. Тонические волокна имеют множественную иннервацию, т.е. на одном волокне по всей его длине располагаются несколько синапсов. Каждый импульс приводит к освобождению только небольшого количества ацетилхолина, поэтому степень деполяризации зависит от частоты стимуляции, а механический ответ носит градуальный характер. Тонические волокна участвуют в сохранении позы. Фазные волокна, наоборот, сокращаются по принципу «все или ничего». Как только нервный импульс достигнет надпороговой силы, происходит генерация распространяющегося потенциала действия, максимальное освобождение Са2+ из везикул и максимальное (одиночное) по силе сокращение, которое уже не возрастает при увеличении интенсивности раздражения. Вместе с тем, при электрическом раздражении целой мышцы (при наложении на нее электродов) сила сокращения мышцы зависит от интенсивности стимула. Стимул, едва превышающий порог, вызовет ответ по принципу «все или ничего» только в ближайших волокнах, где плотность тока максимальна. При этом при небольшой силе раздражения возбуждаются более возбудимые волокна. Для возбуждения всех волокон требуется много большая интенсивность стимула. Причина такого положения в том, что каждое волокно электрически изолировано друг от друга. Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна I типа и «быстрые» – волокна II типа. Различие между быстрыми и медленными волокнами легко заметно даже гистологически: как правило, быстрые волокна содержат мало митохондрий и очень мало миоглобина, поэтому окрашены слабо. В силу этого их часто называют «белыми». Поскольку мощность ресинтеза АТФ в них невелика (преобладает анаэробный гликолиз), то они быстро утомляются, но способны развивать большое напряжение. Быстрые волокна подразделяются как минимум также на две группы. Одна группа – это быстрые гликолитические волокна II B типа, вторая – быстрые окислительные волокна II A типа, способные использовать как окислительное фосфорилирование, так и гликолиз. Волокна типа II А присутствуют у некоторых млекопитающих, но у человека имеются в небольших количествах. Медленные волокна содержат много митохондрий и миоглобина, имеют больше капилляров, вследствие этого имеют выраженное красноватое окрашивание и называются «красными». В силу большой мощности аэробного ресинтеза АТФ в окислительном фосфорилировании утомляются мало, но развивают относительно небольшое напряжение. У млекопитающих и некоторых других видов часть мышц представлена почти исключительно волокнами одного типа. Например, длинный разгибатель пальца включает только быстрые волокна, а камбаловидная мышца – только медленные. У человека же мышцы, включая отмеченные, преимущественно смешанные. Но все же в длинном разгибателе пальца доминируют быстрые, а в камбаловидной мышце медленные волокна. |