Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

ЛЕКЦИЯ 7. МЕХАНИЗМЫ И СПОСОБЫ ТОРМОЖЕНИЯ В ЦНС. КООРДИНАЦИОННАЯ ДЕЯТЕЛЬНОСТЬ ЦНС.7.1. Процессы торможения в ЦНС: механизм постсинаптического и пресинаптического торможений, посттетаническое и пессимальное торможение. Значение торможения.Торможением называется процесс прекращения текущего возбуждения или предотвращение возможного возбуждения нервной структуры. Явление центрального торможения было открыто И.М.Сеченовым в 1862 г. Он обнаружил, что если на поперечный разрез зрительных бугров лягушки наложить кристаллик поваренной соли или подействовать электрическим слабым током, то время рефлекса Тюрка резко удлиняется (рефлекс Тюрка - сгибание лапки при погружении ее у в кислоту). Вскоре были открыты новые факты, демонстрирующие явления торможения в ЦНС. Гольц показал, что рефлекс Тюрка затормаживается при сдавливании пинцетом другой лапки, Шеррингтон доказал наличие торможения рефлекторного сокращения разгибателя при осуществлении сгибательного рефлекса. Было доказано, что при этом интенсивность рефлекторного торможения зависит от соотношения силы возбуждающего и тормозящего раздражителей. История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И. М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции). Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образований. Впоследствии это предположение нашло прямое экспериментальное подтверждение (Экклс, Реншоу), когда было показано существование специальных вставочных нейронов, имеющих синаптические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двигательных нейронов. В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное. В центральной нервной системе существует несколько способов торможения, имеющих разную природу и разную локализацию. но в принципе основанных на одном механизме - увеличении разницы между критическим уровнем деполяризации и величиной мембранного потенциала нейронов. 1. Постсинаптическое торможение. Тормозные нейроны. В настоящее время установлено, что в ЦНС наряду с возбуждающими нейронами существуют и особые тормозные нейроны. Примером может служить т.н. клетка Реншоу в спинном мозге. Реншоу открыл, что аксоны мотонейронов перед выходом из спинного мозга дают одну или несколько коллатералей, которые заканчиваются на особых клетках, чьи аксоны образуют тормозные синапсы на мотонейронах данного сегмента. Благодаря этому возбуждение, возникающее в мотонейроне, по прямому пути распространяется на периферию к скелетной мышце, а по коллатерали активирует тормозную клетку, которая подавляет дальнейшее возбуждение мотонейрона. Это механизм, автоматически охраняющий нервные клетки от чрезмерного возбуждения. Торможение, осуществляющееся при участии клеток Реншоу, получило название возвратного постсинаптического торможения. Тормозным медиатором у клетки Реншоу является глицин. Нервные импульсы, возникающее при возбуждении тормозящих нейронов, не отличаются от потенциалов действия обычных возбуждающих нейронов. Однако в нервных окончаниях тормозящих нейронов под влиянием этого импульса выделяется медиатор, который не деполяризует, а, наоборот, гиперполяризует постсинаптическую мембрану. Эта гиперполяризация регистрируется в форме тормозного постсинаптического потенциала (ТПСП) - электроположительной волны. ТПСП ослабляет возбудительный потенциал и препятствует тем самым достижению критического уровня деполяризации мембраны, необходимого для возникновения распространяющегося возбуждения. Постсинаптическое торможение можно устранить стрихнином, который блокирует тормозные синапсы. 2. Посттетаническое торможение. Особым видом торможения является такое, которое возникает в случае, если после окончания возбуждения в клетке возникает сильная гиперполяризация мембраны. Возбуждающий постсинаптический потенциал в этих условиях оказывается недостаточным для критической деполяризации мембраны, и генерации распространяющегося возбуждения. Причина такого торможения в том, что следовые потенциалы способны к суммации, и после серии частых импульсов возникает суммация положительного следового потенциала. 3.Пессимальное торможение. Торможение деятельности нервной клетки может осуществляться и без участия особых тормозных структур. В этом случае оно возникает в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под влиянием слишком частых импульсов (как пессимум в нервно-мышечном препарате). К пессимальному торможению особо склонны промежуточные нейроны спинного мозга, нейроны ретикулярной формации. При стойкой деполяризации в них наступает состояние, подобное катодической депрессии Вериго.
4. Пресинаптическое торможение. Оно открыто в ЦНС сравнительно недавно, поэтому изучено меньше. Пресинаптическое торможение локализуется в пресинаптических терминалях перед синаптической бляшкой. На пресинаптических терминалях располагаются окончания аксонов других нервных клеток, образующих здесь аксо-аксональные синапсы. Медиаторы их деполяризуют мембрану терминалей и приводят в состояние, подобное катодической депрессии Вериго. Это обусловливает частичную или полную блокаду проведения по нервным волокнам возбуждающих импульсов, идущих к нервным окончаниям. Пресинаптическое торможение обычно длительное. Т  орможение в центральной нервной системе — активный процесс, проявляю-щийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью. Торможение в норме неразрывно связано с возбуждением, является его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению последнего. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и заторможенных зон в центральных нервных структурах. Формирующий эффект тормозного процесса развивается в пространстве и во времени. Торможение — врожденный процесс, постоянно совершенствующийся в течение индивидуальной жизни организма. При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток. орможение в центральной нервной системе — активный процесс, проявляю-щийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью. Торможение в норме неразрывно связано с возбуждением, является его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению последнего. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и заторможенных зон в центральных нервных структурах. Формирующий эффект тормозного процесса развивается в пространстве и во времени. Торможение — врожденный процесс, постоянно совершенствующийся в течение индивидуальной жизни организма. При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток.Описанные способы обеспечивают реализацию нескольких видов безусловного торможения: реципрокного, возвратного, латерального, для реализации которых существуют специальные нервные структуры. Возвратным торможением называют нейронную цепь, в которой тормозные вставочные нейроны действуют на те же нервные клетки, которые их возбуждают. Характерный пример возвратного торможения существует в мотонейронах. Мотонейроны посылают коллатерали к вставочным нейронам, аксоны которых образуют тормозные синапсы на мотонейронах. Такая тормозная цепь называется торможением Реншоу, а тормозной вставочный нейрон клеткой Реншоу В такой цепи усиление возбуждения, поступающего к мышце, усиливает торможение мотонейрона под действием клетки Реншоу. Это пример торможения по принципу обратной связи. 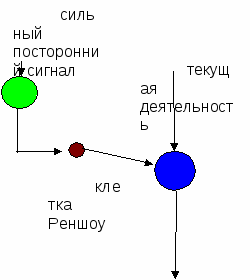 Реципрокное торможение развивается в центрах противоположной деятельности - например, в центрах сгибателей при возбуждении центра разгибателей. Возвратное торможение осуществляет мотонейрон через клетку Реншоу, обеспечивая готовность воспринять новую команду от пирамидной клетки. Реципрокное торможение развивается в центрах противоположной деятельности - например, в центрах сгибателей при возбуждении центра разгибателей. Возвратное торможение осуществляет мотонейрон через клетку Реншоу, обеспечивая готовность воспринять новую команду от пирамидной клетки. Латеральное торможение возникает в центре рефлекса, когда возникает посторонний достаточно сильный сигнал, который через тормозные клетки тормозит текущую деятельность. |