Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
Катаболитная репрессия - это форма позитивного контроля транскрипцией, так как регулирующий белок усиливает транскрипцию оперона. Этот процесс был обнаружен в E.coli и упоминался первоначально как эффект глюкозы, потому что глюкоза подавляла синтез некоторых индуцибeльных ферментов, даже при том, что индуктор присутствовал в окружающей среде. Открытие было сделано процессе изучения регуляции lac-оперона у E.coli. Так как глюкоза разлагается конститутивными ферментами, а лактоза - первоначально индуцибeльными, что случилось, если бы бактерия была выращена в условиях ограниченных количеств глюкозы и лактозы? График роста бактерий в этих условиях будет пред- 115 ставлен двухфазной кривой роста, которая покажет две различные стадии активного роста (рис. 20). течение первой стадии экспоненциального роста бактерии исполь-зуют глюкозу как источник энергии, пока вся глюкоза не израсходуется. Затем после второй лаг-фазы лактоза используется в течение второй фазы экспоненциального роста. В течение периода использования глюкозы лак-тоза не потребляется, потому что клетки неспособны транспортировать и расщеплять дисахарид (лактозу). Глюкоза всегда усваивается первой в предпочтении любому другому сахару. Только после того, как глюкоза полностью израсходуется, будет использоваться лактоза. Лактозный опе-рон будет подавляться даже в тех случаях, когда лактоза (индуктор) при-сутствует в питательной среде. Экологическое объяснение этого процесса состоит в том, что глюкоза - более экономичный источник энергии, чем лактоза, так как ее использование требует меньше ферментов, чем утили-зация лактозы. 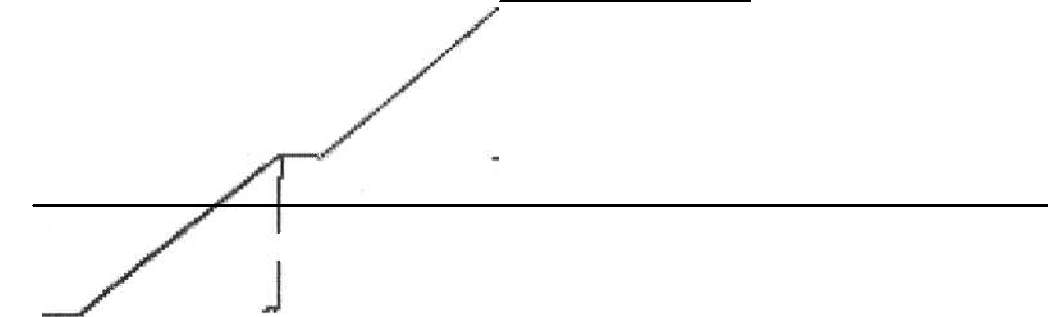 1 7" Утилизация лактозы Оптическая ПЛОТНОСТЬ * Время (часы) Рис. 20. Двухфазный рост. Кривая роста E.coli в условиях лимитирующих концентраций в смеси глюкозы и лактозы Вторичная лаг-фаза в течение двухфазного роста представляет время, требуемое для полной индукции lac-оперона и синтеза ферментов, необхо-димых для использования лактозы (галактозидпермеазы и Р-галактозидазы). Только тогда бактериальный рост происходит за счет лактозы. Глюкоза, как известно, подавляет большое количество индуцибeльных ферментов в клетках бактерий многих видов. Она также подавляет образо-вание индуцибeльных оперонов, запрещая синтез циклического АМФ 116 (цАМФ), нуклеотида, который требуется для инициирования транскрип-ции большого количества индуцибeльных систем фермента, включая lac-оперон. Роль цАМФ сложна. Он обязан активизировать аллостерический белок, названный САР (катаболитный белок-активатор), который связыва-ется с промотором САР и способствует прикреплению РНК-полимеразы (РНКп) к промотору для инициации транскрипции. Таким образом, для инициации транскрипции гена lac-оперона необходима лактоза, чтобы инактивировать lac-ген-репрессор, а также цАМФ, чтобы связаться с САР, который соединяется с ДНК для облегчения транскрипции. Эффект глюкозы выполняет полезную функцию у бактерий, необхо-димую для того, чтобы клетки использовали наиболее доступный источник энергии. Для многих бактерий глюкоза - самый обычный и хорошо утили-зируемый субстрат для роста. То есть она косвенно запрещает синтез фер-ментов, для того чтобы усваивались более бедные источники энергии. В присутствии глюкозы деятельность аденилатциклазы (AЦ) блокирована. AЦ обязана синтезировать цАМФ из ATФ. Поэтому, если концентрация цАМФ низкая, САР неактивен и транскрипция не происходит. В отсутст-вии глюкозы (концентрация цАМФ высокая) САР активизируется цАМФ, транскрипция происходит (в присутствии лактозы). Многие положитель-но управляемые промоторы, типа lac-промотора, не полностью функцио-нируют в присутствии одной РНКп и требуют активации САРом, который кодируется отдельным регулирующим геном и присутствует в конститу-тивных уровнях. САР активен только в присутствии цАМФ. Прикрепление цАМФ к САРу вызывает конформационное изменение в белке, что позво-ляет ему связаться с промотором рядом с РНКп связывающим сайтом. САР может очевидно взаимодействовать с РНКп и тем самым усиливать транс-крипцию оперона примерно в 50 раз. Позитивная регуляция работы lac-оперона представлена на рис. 21. основе другого феномена - феномена репрессии - лежит тот же принцип регуляции. Однако в репрессируемой системе ген-регулятор кон-тролирует синтез апорепрессора, то есть неактивного репрессора, который также имеет два активных центра: один - для взаимодействия с метаболи-том (корепрессором), а другой - для специфического связывания с геном-оператором. Апорепрессор становится активным и подавляет работу опе-рона лишь после взаимодействия с соответствующим корепрессором (ме-таболитом). 117 Типичным примером репрессируемой системы является система син-теза ферментов пути образования триптофана у E.coli (рис. 22). отсутствии триптофана апорепрессор неактивен и не блокирует работу трипто фанов ого оперона. Циклический АМФ  А Q>> —■ Транскрипция
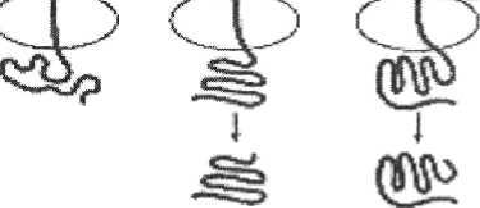 Рис. 21. Катаболитная репрессия - позитивная регуляция lac-оперона. Действие - усиление транскрипции. Белок САР активизируется цАМФ для связывания с lac-опероном и облегчает прикрепление РНКп к промотору для транскрипции генов, кодирующих ферменты утилизации лактозы Регулиорные гены Структурные гены 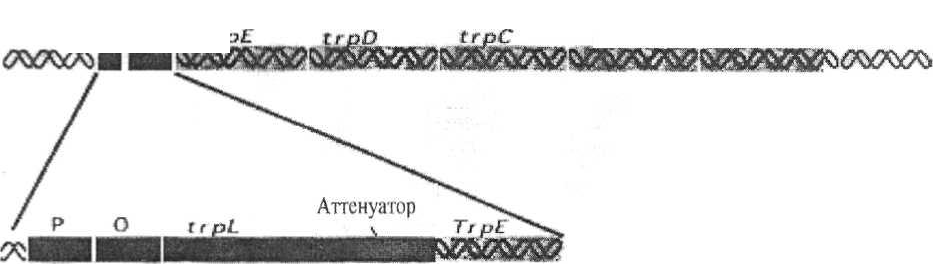 ttpB Промотср Рис. 22. Генетическая организация оперона триптофана и элементы его регуляции При избыточном содержании триптофана в среде, в которой размно-жается E. coli, он, выполняя роль корепрессора, связывается с апорепрессо- ром и вызывает его аллостерическое превращение в активный репрессор. 118 Последний связывается с геном-оператором, что и приводит к пре-кращению дальнейшей транскрипции структурных цистронов этого опе-рона и подавлению синтеза ферментов. Особенностью триптофанового оперона является наличие в нем ме-жду промоторно-операторным участком и его первым структурным цис-троном особой последовательности приблизительно из 150 пар нуклеоти-дов, получившей название лидерной последовательности, или аттенуатора. Роль аттенуатора состоит в регуляции активности РНК-полимеразы и за-ключается в том, что даже при незначительном избытке триптофана в клетке транскрипция оперона большинством молекул РНК-полимераз преждевременно обрывается в области аттенуатора. По мере же снижения концентрации триптофана все больше и больше молекул РНК-полимераз «проскакивают» этот участок и становятся способными транскрибировать весь оперон. Наоборот, при большом избытке триптофана его молекулы переводят апорепрессор в корепрессор и транскрипция оперона подавляет-ся (рис. 23). Аттенуатор 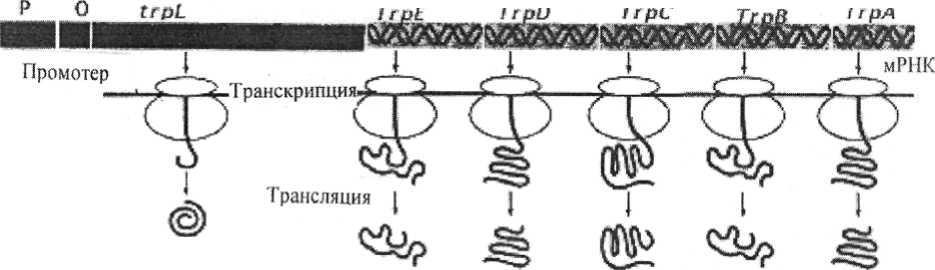
Рис. 23. Репрессия работы оперона триптофана. В отсутствии триптофана неактивный репрессор не может связаться с опероном, чтобы блокировать транскрипцию. Клетки должны синтезировать аминокислоту Помимо негативных, существуют и позитивные механизмы контроля выражения генетической информации. Они были обнаружены при изуче-нии арабинозного оперона у E.coli. Система позитивного контроля являет-ся необходимым атрибутом координированного управления различными 119 оперонами. Таким образом, благодаря сочетанию механизмов индукции и репрессии, негативного и позитивного контроля выражения генетической информации обеспечивается определенная координация между различны-ми функциональными группами оперонов. 4.5. Механизмы изменчивости бактерий. Роль внехромосомных факторов наследственности и транспонируемых элементов Первое, что поразило исследователей, когда они поближе познако-мились с миром прокариот, - огромное разнообразие присущих им при-знаков. В процессе экспериментальной работы с прокариотами исследова-тели часто наблюдали, что популяция одного вида при культивировании в течение длительного времени или в разных условиях подвержена измене-ниям. Накопилось огромное количество результатов, иллюстрирующих эти изменения, однако механизмы, лежащие в основе наблюдаемых явлений, были непонятны. Успехи, достигнутые за последние десятилетия XX в. в области изучения строения и функционирования генетического аппарата прокариот, позволили разобраться в этом вопросе. Прежде чем переходить дальнейшему изложению материала, целесообразно ввести некоторые понятия. Генотипом, или геномом, называют совокупность всех генов,при-сущих данному организму, то есть его генетическую конституцию. Под фенотипом понимают совокупность признаков, присущих дан-ному организму. Оказалось, что все наблюдаемые изменения можно разде-лить на два типа. К первому относят те из них, которые, как правило, про-являются у подавляющего большинства особей в популяции при измене-нии внешних условий и наблюдаются до тех пор, пока действует фактор, вызвавший эти изменения. Такой тип изменчивости получил название не-наследственного, или модификационного, а само явление названо моди-фикацией. Ко второму типу относятся изменения признаков,которые пер-воначально возникают как редкие события в популяции особей (с частотой 1 на 104 - 1011 клеток). Если измененные особи имеют некоторое преиму-щество перед неизмененными, выражающееся в повышенной скорости роста или жизнеспособности, они постепенно накапливаются в популяции и вытесняют исходные особи. Изучение особенностей второго типа изме-нений привело к заключению, что последние возникают случайно. И нако- 120 нец, эти изменения постоянны, то есть передаются из поколения в поколе-ние при размножении организма. Такой тип изменчивости был назван на-следств енным. МОДИФИКАЦИЯ Это изменение, происходящее на уровне фенотипа и не затрагиваю-щее клеточный генотип. Все признаки клетки определяются ее генотипом, но в определенных условиях она пользуется не всей заложенной в ней ге-нетической информацией, количество которой гораздо больше, чем необ-ходимо клетке для существования в конкретных условиях. Реакция клетки на изменение внешних условий приводит к проявлению каких-то новых признаков, свойств, которые не обнаруживались в исходной культуре. Од-нако информация, необходимая для проявления этих признаков, обяза-тельно содержится в клеточном геноме. Модификация есть результат пла-стичности клеточного метаболизма, приводящего к фенотипическому про-явлению «молчащих» генов в конкретных условиях. Таким образом, мо-дификационные изменения имеют место в рамках неизменного клеточного генотипа. Существует несколько типов модификационных изменений. Наибо-лее известны адаптивные модификации, то есть ненаследственные измене-ния, полезные для организма и способствующие его выживанию в изме-нившихся условиях. Причины адаптивных модификаций кроются в меха-низмах регуляции действия генов. Адаптивной модификацией является адаптация клеток Е.соli к лактозе как к новому субстрату. У ряда бактерий обнаружена универсальная адаптивная реакция в ответ на различные стрессовые воздействия (высокие и низкие температуры, резкий сдвиг рН др.), проявляющаяся в интенсивном синтезе небольшой группы сходных белков. Такие белки получили название белков теплового шока, а само яв-ление - синдром теплового шока. Еще не ясны те регуляторные механиз-мы, которые запускаются в клетке при воздействиях, вызывающих син-дром теплового шока, но очевидно, что это универсальный механизм не-специфических адаптивных модификаций. Не все модификации обяза-тельно адаптивны. Таким образом, модификационная изменчивость не затрагивает ге-нетической конституции организма, то есть не является наследственной. В то же время она вносит определенный вклад в процесс эволюции. Адап- тивные модификации расширяют возможности организма к выживанию и 121 размножению в более широком диапазоне условий внешней среды. Возни-кающие в этих условиях наследственные изменения подхватываются есте-ственным отбором и таким путем происходит более активное освоение но-вых экологических ниш и достигается более эффективная приспособляе-мость к ним. Наследственные изменения Можно подразделить на изменения, возникающие в результате мута-ций и рекомбинаций генетического материала. МУТАЦИИ Скачкообразные изменения в генетическом материале клетки, при-водящие к появлению новых признаков, получили название мутаций. Они возникают в популяции особей всегда, часто без видимых воздействий на популяцию. Такие мутации, причины возникновения которых нам неиз-вестны, называются спонтанными. Повышать частоту мутаций по сравне-нию с фоном, то есть индуцировать их, могут физические, химические и биологические факторы, действующие на генетический материал клетки. Физические факторы - это прежде всего коротковолновое излучение (ультрафиолетовые и рентгеновские лучи). Получение индуцированных мутаций (мутантов) - один из основных способов изучения генетики мик-роорганизмов. химическим мутагенам относят аналоги оснований, акридины, ал-килирующие и дезаминирующие агенты. Биологические факторы - это, в первую очередь мигрирующие элементы (транспозоны и IS-элементы). Мутации, независимо от того, имеют ли они спонтанное происхож-дение или индуцированы каким-либо мутагеном, по характеру перестроек, происшедших в ДНК, можно разделить на мутации, состоящие в измене-нии одного нуклеотидного остатка молекулы ДНК, так называемые точко-вые мутации, и мутации, при которых наблюдается изменение участка мо-лекулы ДНК размером больше одного нуклеотида. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||