Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
хромосомы. Полуконсервативный механизм репликации ДНК (рис. 15), в резуль-тате которого из одной родительской двухцепочной молекулы образуются две дочерние молекулы, содержащие по одной родительской и одной вновь синтезированной комплементарной полинуклеотидной цепи, наи-лучшим образом обеспечивает идентичность исходной и синтезированных молекул и, следовательно, сохранность видоспецифической наследствен-ной информации в ряду поколений клеток и организмов. Частота ошибок, возникающих в процессе репликации, порядка 10-7. Реализация наследственной информации в процессе жизненного цикла (онтогенеза) организма - двухступенчатый процесс. Сначала с опре-деленных участков ДНК информация переписывается (транскрибируется) виде комплементарных нуклеотидных последовательностей молекул иРНК, которая перемещается в цитоплазму, связывается с рибосомами, и в рибосоме с иРНК осуществляется перевод (трансляция) генетической ин-формации в определенную последовательность аминокислотных остатков молекулы белка. Процесс транскрипции находится в клетке под строгим контролем, поэтому имеет место как неодинаковое транскрибирование во времени разных участков ДНК (генов), так и неодинаковая скорость, с ко-торой гены могут транскрибироваться. В результате количество молекул 105 иРНК в клетке, комплементарных разным генам, сильно различается. Хотя целом механизмы синтеза ДНК и РНК сходны, процесс транскрипции не обладает той степенью точности, которая характерна для репликации ДНК. Однако поскольку иРНК не способна к самовоспроизведению, возникаю-щие при ее синтезе ошибки в последующих клеточных генерациях не вос-производятся и, следовательно, не могут наследоваться. / \ \ Лояцкоысврватавныи Дцслерриеньги Консервативный ( те- конец) Рис. 15. Полуконсервативный, дисперсивный и консервативный (конец-в-конец) способы распределения двух родительских цепей ДНК Несмотря на то, что механизм трансляции отличается высокой точ-ностью, вероятность ошибки в целом выше, чем в случае синтеза молекул нуклеиновых кислот и РНК. Наиболее уязвимый этап - «узнавание» с по-мощью фермента аминокислоты соответствующей молекулой тРНК. Час-тота возникновения ошибок на этом этапе около 10-4 , что и определяет, 106 возможно, уровень точности процесса синтеза белка в целом. Однако, как в случае синтеза РНК, белки в процессе трансляции, приводящие к син-тезу измененной молекулы белка, не воспроизводятся, если они не закоди-рованы исходно в генетическом материале. Только изменения, происходя-щие в молекулах ДНК, могут сохраняться в ряду поколений, поскольку они воспроизводятся в процессе репликации. многих бактерий обнаружены нехромосомные генетические эле-менты: плазмиды, умеренные фаги, мигрирующие элементы транспозоны IS-элементы (от англ. insertion sequences - вставные последовательности). Таким образом, у прокариот большой объем генетической информации оказывается рассредоточенным в нехромосомных элементах. Это заставля-ет по-новому подходить к вопросу об организации генетической информа-ции в мире прокариот. Особенность генетической информации, содержащейся в нехромо-сомных элементах, - ее необязательность для жизнедеятельности бакте-рий, то есть в ее отсутствии бактериальная клетка жизнеспособна, но, как видно из дальнейшего материала, важная роль нехромосомных генетиче-ских элементов заключается в том, что они расширяют возможности суще-ствования бактериального вида, обеспечивают обмен генетическим мате-риалом на большие расстояния по горизонтали и играют определенную роль в эволюции прокариот. бактерий различают три типа репликации ДНК: вегетативную, конъюгативную и репаративную. Вегетативная репликация хромосомной и плазмидной ДНК обу-словливает передачу генетической информации по вертикали, то есть по наследству от родительской клетки дочерним. Она контролируется соот-ветственно хромосомными и плазмидными генами. Конъюгативная репликация осуществляется при конъюгативномспособе обмена генетическим материалом и контролируется только плаз-мидными генами. При ней происходит достройка нити ДНК, комплемен-тарной нити, передаваемой от донора реципиенту. Репаративная репликация является механизмом,посредством ко-торого происходит устранение из ДНК структурных повреждений, или за-ключительный этап генетической рекомбинации. Эти процессы контроли-руются хромосомными и плазмидными генами. 107 Вегетативная репликация. Репликация хромосомной ДНК осуще-ствляется от начальной точки ori (от англ. origin - начало), носит полукон-сервативный характер (см. рис. 15), идет одновременно в двух направлени-ях и заканчивается в точке terminus. Хромосома бактериальной клетки свя-зана в области ori с цитоплазматической мембраной. Репликация ДНК ка-тализируется ДНК-полимеразами. Сначала происходит раскручивание (деспирализация) двойной цепи ДНК, в результате чего образуется репликативная вилка (разветвленные цепи) (рис. 16). Одна из цепей, достраиваясь, связывает нуклеотиды от 5' - 3'-концу, другая - достраивается посегментно. \ 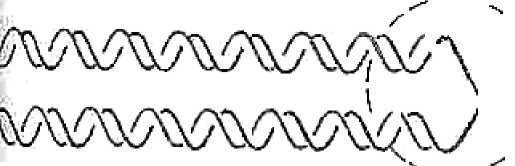 Рвплицированная (дочернин) 'ч ' (родт -елъскэя) ДНК ДНК Рис. 16. Репликативная вилка Поскольку цепи ДНК антипараллельны (если одна нить начинается с 5'-конца, другая - с 3'-конца), а ДНК-полимераза III осуществляет синтез ДНК только в направлении 5'>3', репликация происходит своеобразно (рис. 17): на одной из расплетенных нитей - «прямой», «лидерной» или ве-дущей - она идет непрерывно, а на другой - «отстающей» - ДНК-полимераза III должна возвращаться, чтобы наращивать нить тоже в на-правлении 5'>3' прерывисто, через образование сегментов Оказаки, у бак-терий длиной около 1000 нуклеотидов (у эукариот - около 200 - 300 нук-леотидов). Синтез каждого сегмента Оказаки происходит последовательно через следующие стадии: Раскручивание нитей ДНК. Расплетение (разделение нитей). Стабилизация однонитевых участков. Формирование праймосомы (праймосома - мультиферментный комплекс, в который входят фермент ДНК-праймаза и ряд других белков). Синтез затравочной РНК с участием ДНК-праймазы (от англ. prime - подготавливать). Затравочная РНК необходима для синтеза каждо- 108 го сегмента Оказаки потому, что сама ДНК-полимераза не способна ини-циировать синтез ДНК, для этого ей нужна специальная затравка, роль ко-торой и выполняют короткие, длиной не более 10 нуклеотидов, фрагменты РНК, комплементарные ДНК-матрице. 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||