Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
70 Аспарагин используется только для синтеза белковых молекул. Во все остальные аминокислоты азот вводится посредством реакций переами-нирования, катализируемых соответствующими аминотрансферазами, при этом во всех реакциях одним из участников является глутаминовая кислота: глутаминовая кислота + щавелевоуксусная кислота —> —> аспарагиновая кислота + а-кетоглутаровая кислота. Еще одним путем включения азота аммиака в состав органических со-единений является реакция, приводящая к образованию карбамоилфосфата: NН3 + СО2 + 2АТФ -»Н2N — СО — О — РО3Н2 + 2АДФ + ФH. Карбамоилфосфат Дальнейшее использование азота карбамоилфосфата происходит по двум направлениям: для синтеза пиримидинов и аргинина. Биосинтез мононуклеотидов. Из мононуклеотидов построены нук-леиновые кислоты (РНК, ДНК) клеток. Кроме того, мононуклеотиды вхо-дят в состав многих коферментов и участвуют, таким образом, в осуществ-лении различных каталитических функций. Центральное место в биосин-тезе мононуклеотидов занимает синтез пуриновых и пиримидиновых азо-тистых оснований. Большинство прокариот способно к синтезу этих со-единений из низкомолекулярных предшественников. Синтез пуриновых и пиримидиновых мононуклеотидов осуществля-ется независимыми путями. результате последовательных ферментативных реакций при синте-зе пуриновых нуклеотидов образуется инозиновая кислота, из которой пу-тем химических модификаций пуринового кольца синтезируются аденило-вая (АМФ) и гуаниловая (ГМФ) кислоты. Первым синтезируемым пиримидиновым нуклеотидом является оро-тидиловая кислота, декарбоксилирование которой приводит к образованию уридиловой кислоты (УМФ). Последняя служит предшественником цити-диловых нуклеотидов, но соответствующее превращение происходит толь-ко на уровне трифосфатов, поэтому сначала из УМФ образуется УТФ, аминирование которого приводит к возникновению ЦТФ. Дезоксирибо-нуклеотиды образуются в результате восстановления соответствующих рибонуклеотидов на уровне дифосфатов (для некоторых прокариот описа-но подобное превращение на уровне трифосфатов). Синтез специфическо- 71 го для ДНК нуклеотида - тимидиловой кислоты - происходит путем фер-ментативного метилирования дезоксиуридиловой кислоты. Многие прокариоты способны использовать содержащиеся в пита-тельной среде готовые пуриновые и пиримидиновые основания, их нук-леозиды и нуклеотиды, имея ферменты, катализирующие следующие эта-пы взаимопревращений экзогенных пуриновых и пиримидиновых произ-водных: азотистое основание <-> нуклеозид <-> нуклеотид (моно ■^•ди <-> трифосфат). МЕХАНИЗМЫ ПИТАНИЯ Поступление различных веществ в бактериальную клетку зависит от величины и растворимости их молекул в липидах или воде, рН среды, кон-центрации веществ, различных факторов проницаемости мембран и др. Клеточная стенка пропускает небольшие молекулы и ионы, задерживая макромолекулы массой более 600 Да. Основным регулятором поступления веществ в клетку является цитоплазматическая мембрана. Условно можно выделить четыре механизма проникновения пита-тельных веществ в бактериальную клетку: простая диффузия, облегченная диффузия, активный транспорт, транслокация групп. Наиболее простой механизм поступления веществ в клетку - про-стая диффузия, при которой перемещение веществ происходит вследст-вие разницы их концентрации по обе стороны цитоплазматической мем-браны. Вещества проходят через липидную часть цитоплазматической мембраны (органические молекулы, лекарственные препараты) и реже по заполненным водой каналам в цитоплазматической мембране. Пассивная диффузия осуществляется без затраты энергии. Облегченная диффузия происходит также в результате разницыконцентрации веществ по обе стороны цитоплазматической мембраны. Однако этот процесс осуществляется с помощью молекул-переносчиков, локализующихся в цитоплазматической мембране и обладающих специ-фичностью. Каждый переносчик транспортирует через мембрану соответ-ствующее вещество или передает другому компоненту цитоплазматиче-ской мембраны - собственно переносчику. Белками-переносчиками могут быть пермеазы, место синтеза которых - цитоплазматическая мембрана. Облегченная диффузия протекает без затраты энергии, вещества переме-щаются из области с более высокой концентрацией в область с более низкой. 72 Активный транспорт происходит с помощью пермеаз и направленна перенос веществ меньшей концентрации в сторону большей, то есть как бы против течения, поэтому данный процесс сопровождается затратой ме-таболической энергии (АТФ), образующейся в результате окислительно-восстановительных реакций в клетке. Перенос (транслокация) групп сходен с активным транспортом,ноотличается тем, что переносимая молекула видоизменяется в процессе пе-реноса, например фосфорилируется. Выход веществ из клетки осуществляется за счет диффузии и при участии транспортных систем. ФЕРМЕНТЫ БАКТЕРИЙ Ферменты распознают соответствующие метаболиты (субстраты), вступают с ними во взаимодействие и ускоряют химические реакции. Яв-ляются белками, участвуют в процессах анаболизма (синтеза) и катабо-лизма (распада), то есть метаболизма. Многие ферменты взаимосвязаны со структурами микробной клетки. Например, в цитоплазматической мембра-не имеются окислительно-восстановительные ферменты, участвующие в дыхании и делении клетки. Окислительно-восстановительные ферменты цитоплазматической мембраны и ее производных обеспечивают энергией интенсивные процессы биосинтеза различных структур, в том числе кле-точной стенки. Ферменты, связанные с делением и аутолизом клетки, об-наруживаются в клеточной стенке. Так называемые эндоферменты ката-лизируют метаболизм, проходящий внутри клетки. Экзоферменты выде-ляются клеткой в окружающую среду, расщепляя макромолекулы пита-тельных субстратов до простых соединений, усваиваемых клеткой в каче-стве источников энергии, углерода и др. Некоторые экзоферменты (пени-циллиназа и др.) инактивируют антибиотики, выполняя защитную функ-цию. Различают конститутивные и индуцибельные ферменты. Консти-тутивные ферменты синтезируются клеткой непрерывно,вне зависимо-сти от наличия субстратов в питательной среде. Индуцибельные (адаптив-ные) ферменты синтезируются только при наличии в среде субстрата данного фермента. Например, кишечная палочка на среде с глюкозой практически не образует Р-галактозидазу, но резко увеличивает ее синтез при выращивании на среде с лактозой или другим Р-галактозидом. 73 Некоторые ферменты (так называемые ферменты агрессии) разру-шают ткань и клетки, обусловливая широкое распространение в инфици-рованной ткани микроорганизмов и их токсинов. К таким ферментам отно-сят гиалуронидазу, коллагеназу, дезоксирибонуклеазу, нейраминидазу, ле-цитовителлазу и др. Так, гиалуронидаза стрептококков, расщепляя гиалу-роновую кислоту соединительной ткани, способствует распространению стрептококков и их токсинов. Известно более 2000 ферментов. Они объединены в шесть классов: оксидоредуктазы - окислительно-восстановительные ферменты (к ним от-носятся дегидрогеназы, оксидазы и др.); трансферазы, переносящие от-дельные радикалы и атомы от одних соединений к другим; гидролазы, ус-коряющие реакции гидролиза, то есть расщепления веществ на более про-стые с присоединением молекул воды (эстеразы, фосфатазы, глюкозидазы др.); лиазы, отщепляющие от субстратов химические группы негидроли-тическим путем (карбоксилазы и др.); изомеразы, превращающие органи-ческие соединения в их изомеры (фосфогексоизомераза и др.); лигазы, или синтетазы, ускоряющие синтез сложных соединений из более простых (ас-парагинсинтетаза, глютаминсинтетаза и др.). Различия в ферментном составе используются для идентификации микроорганизмов, так как они определяют их различные биохимические свойства: сахаролитические (расщепление сахаров), протеолитические (разложение белков) и другие, выявляемые по конечным продуктам рас-щепления (образование щелочей, кислот, сероводорода, аммиака и др.). Ферменты микроорганизмов используют в генетической инженерии (рестриктазы, лигазы и др.) для получения биологически активных соеди-нений, уксусной, молочной, лимонной и других кислот, молочнокислых продуктов, в виноделии и других отраслях. Ферменты применяют в каче-стве биодобавок в стиральные порошки для уничтожения загрязнений бел-ковой природы. 3.3. Дыхание прокариот (энергетический метаболизм) Энергетический метаболизм (катаболизм) - это поток реакций, со-провождающихся мобилизацией энергии и преобразованием ее в электро-химическую (А(Хн+) или химическую (АТФ) форму, которая затем может быть использована во всех энергозависимых процессах. 74 Существуют группы прокариот, энергетический метаболизм которых не связан с превращениями органических соединений (прокариоты с фото-лито- и хемолитотрофным типом энергетического обмена). По отношению такого рода энергетическим процессам термин «катаболизм» неприме-ним. У них функционирует только один поток превращений органических соединений углерода - анаболический. Энергетические процессы прокариот по своему объему (масштабно-сти) значительно превосходят процессы биосинтетические, а их протека-ние приводит к существенным изменениям в окружающей среде. Разнооб-разны и необычны в этом отношении возможности прокариот, способы их энергетического существования. Все это вместе взятое сосредоточило внимание исследователей в первую очередь на изучении энергетического метаболизма прокариот. Энергетические ресурсы. Организмы могут использовать не все ви-ды энергии, существующей в природе. Недоступными для них являются ядерная, механическая, тепловая виды энергии. Чтобы теплота могла слу-жить источником энергии, необходим большой перепад температур, кото-рый в живых организмах невозможен. Доступными для живых систем внешними источниками энергии (энергетическими ресурсами) являются электромагнитная (физическая) энергия (свет определенной длины волны) и химическая (восстановленные химические соединения). Способностью использовать энергию света обладает большая группа фотосинтезирующих организмов, в том числе и прокариот, имеющих фоторецепторные молеку-лы нескольких типов (хлорофиллы, каротиноиды, фикобилипротеины). Для всех остальных организмов источниками энергии служат процессы окисления химических соединений. Часто энергетическими ресурсами служат биополимеры, находящиеся в окружающей среде (полисахариды, белки, нуклеиновые кислоты), а также липиды. Прежде чем быть исполь-зованными, биополимеры должны быть гидролизованы до составляющих их мономерных единиц. Этот этап весьма важен по следующим причинам. Белки и нуклеиновые кислоты отличаются исключительным разнообрази-ем. Количество видов белков исчисляется тысячами, после гидролиза же образуется только 20 аминокислот. Все разнообразие нуклеиновых кислот (ДНК и РНК) после гидролиза сводится к 5 видам нуклеотидов. Таким об-разом, расщепление полимеров до мономерных единиц резко сокращает набор химических молекул, которые могут быть использованы организмом. 75 Полимерные молекулы расщепляются до мономеров с помощью ферментов, синтезируемых и выделяемых прокариотами в окружающую среду (экзоферментов). Крахмал и гликоген гидролизуются амилазами, гликозидные связи целлюлозы расщепляются целлюлазой. Многие бакте-рии образуют пектиназу, хитиназу, агаразу и другие ферменты, гидроли-зующие соответствующие полисахариды и их производные. Белки расще-пляются внеклеточными протеазами, воздействующими на пептидные свя-зи. Нуклеиновые кислоты гидролизуются рибо- и дезоксирибонуклеазами. Образующиеся небольшие молекулы легко транспортируются в клетку че-рез мембрану. Процесс распада жирных кислот локализован в клетке и включает несколько этапов. На первом из них жирная кислота с помощью соответствующего фермента превращается в КоА-производное, которое окисляется в Р-положении с последующим отщеплением ацетил-КоА. Дру-гим продуктом реакции является КоА-производное жирной кислоты, уко-роченное на два углеродных атома. Ацетил-КоА по катаболическим кана-лам используется для получения клеткой энергии. Процесс расщепления биополимеров не связан с образованием сво-бодной, то есть доступной клетке, энергии. Происходящее при этом рас-сеивание энергии также невелико. Образовавшиеся мономеры подвергают-ся в клетке дальнейшим ферментативным превращениям, в результате ко-торых путем перестройки химической структуры возникают молекулы, включающиеся на каком-либо этапе в качестве метаболитов в функциони-рующие клеточные катаболические системы. Основные из них: путь Эм-бдена - Мейергофа - Парнаса (гликолиз), окислительный пентозофосфат-ный путь, путь Энтнера - Дудорова и цикл трикарбоновых кислот (ЦТК). Общее для всех катаболических путей - многоступенчатость процесса окисления исходного субстрата. На некоторых этапах окисление субстрата сопряжено с образованием энергии в определенной форме, в которой эта энергия может использоваться в самых разнообразных энергозависимых процессах. Таким образом, внешние доступные организмам источники энергии (свет, химические соединения) должны быть трансформированы в клетке в определенную форму, чтобы обеспечить внутриклеточные потребности в энергии. 76 Дыхание бактерий. Дыхание, или биологическое окисление, осно-вано на окислительно-восстановительных реакциях, идущих с образовани-ем АТФ-универсального аккумулятора химической энергии. Энергия не-обходима микробной клетке для ее жизнедеятельности. При дыхании про-исходят процессы окисления и восстановления: окисление - отдача доно-рами (молекулами или атомами) водорода или электронов; восстановление - присоединение водорода или электронов к акцептору. Акцептором водо-рода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат (такое дыхание назы-вается анаэробным - нитратным, сульфатным, фумаратным). Анаэробиоз (от греч. aer - воздух, bios - жизнь) - жизнедеятельность, протекающая при отсутствии свободного кислорода. Если донорами и акцепторами во-дорода являются органические соединения, то такой процесс называется брожением. При брожении происходит ферментативное расщепление ор-ганических соединений, преимущественно углеводов, в анаэробных усло-виях. С учетом конечного продукта расщепления углеводов различают спиртовое, молочнокислое, уксусное и другие виды брожения. По отношению к молекулярному кислороду бактерии можно разде-лить на три основные группы: облигатные, то есть обязательные, аэробы, облигатные анаэробы и факультативные анаэробы. Облигатные аэробы -это прокариоты,для роста которых кислороднеобходим. К ним относится большинство прокариотных организмов. Облигатные анаэробы (клостридии ботулизма,газовой гангрены,столбняка, бактероиды и др.) растут только на среде без кислорода, кото-рый для них токсичен. При наличии кислорода бактерии образуют пере-кисные радикалы кислорода, в том числе перекись водорода и супероксид-анион кислорода, токсичные для облигатных анаэробных бактерий, по-скольку они не образуют соответствующие инактивирующие ферменты. Аэробные бактерии инактивируют перекись водорода и супероксид-анион кислорода соответствующими ферментами (каталазой, пероксидазой и су-пероксиддисмутазой). Факультативные анаэробы могут расти как при наличии,так и приотсутствии кислорода, поскольку они способны переключаться с дыхания присутствии молекулярного кислорода на брожение в его отсутствие. Они способны осуществлять анаэробное дыхание, называемое нитратным: 77 нитрат, являющийся акцептором водорода, восстанавливается до молеку-лярного азота и аммиака. Среди облигатных анаэробов различают аэротолерантные бактерии, которые сохраняются при наличии молекулярного кислорода, но не ис-пользуют его. Для выращивания анаэробов в бактериологических лабораториях применяют анаэростаты - специальные емкости, в которых воздух заменя-ется смесью газов, не содержащих кислорода. Воздух можно удалять из питательных сред путем кипячения, с помощью химических адсорбентов кислорода, помещаемых в анаэростаты или другие емкости с посевами. 3.4. Закономерности роста и размножения бактерий РОСТ И РАЗМНОЖЕНИЕ БАКТЕРИЙ Жизнедеятельность бактерий характеризуется ростом - формирова-нием структурно-функциональных компонентов клетки и увеличением са-мой бактериальной клетки, а также размножением - самовоспроизведени-ем, приводящим к увеличению количества бактериальных клеток в попу-ляции. Различают рост клеток и рост популяции. Каждый из них характе-ризуется своими особенностями и закономерностями. Под ростом индиви-дуальной клетки понимают увеличение ее биомассы, наступающее в ре-зультате синтеза клеточного материала. Объем клетки можно вычислить, если известны ее продольные и поперечные размеры. Для шаровидных клеток он определяется по формуле V=4/3nr\ а для цилиндрических - по формуле V= лаг1, где r - радиус клетки, а - длина клетки. Бактерии размножаются путем бинарного деления пополам, реже пу-тем почкования. Актиномицеты, как и грибы, могут размножаться спора-ми, так как являются ветвящимися бактериями, а также путем фрагмента-ции нитевидных клеток. Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а грам - 78 отрицательные - путем перетяжки, в результате чего образуются две оди-наковые клетки (рис. 9).  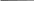   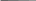 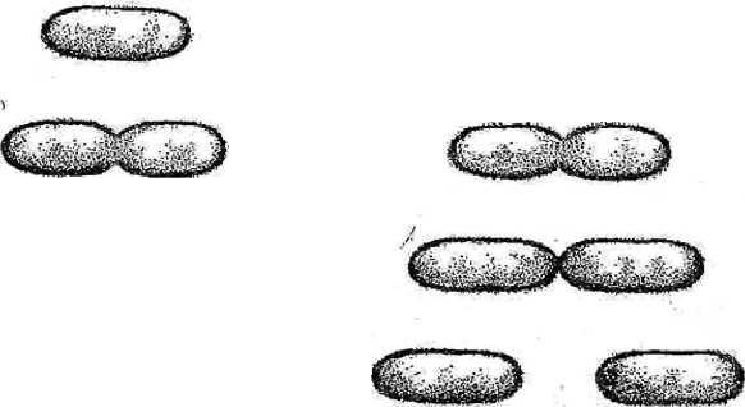 Рис. 9. Процесс роста и размножения бактериальной клетки. Время ге-нерации g представляет собой время между моментом образования бактерии и моментом, когда она расщепляется на две дочерние клетки Делению клеток предшествует репликация бактериальной хромосо-мы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается каждая нить достраивается комплементарной нитью), приводящая к уд-воению молекул ДНК бактериального ядра - нуклеоида. Образовавшиеся в результате репликации две хромосомы расходят-ся, чему способствует увеличение размеров растущей клетки: прикреплен-ные к цитоплазматической мембране или ее производным (например, ме-зосомам) хромосомы по мере увеличения объема клетки удаляются друг от друга. Окончательное их обособление завершается образованием перетяж-ки или перегородки деления (рис. 10). Клетки с перегородкой деления расходятся в результате действия ау-толитических ферментов, разрушающих сердцевину перегородки деления. Аутолиз при этом может происходить неравномерно: делящиеся клетки в одном участке остаются связанными частью клеточной стенки в области 79 перегородки деления. Такие клетки располагаются под углом друг к другу, что характерно для дифтерийных коринебактерий. 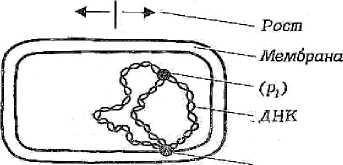 Прикрепление (рг )  S Рис. 10. Механизм распределения бактериальных хромосом КУЛЬТИВИРОВАНИЕ МИКРООРГАНИЗМОВ |