Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
33 КЛЕТОЧНАЯ СТЕНКА Это важный и обязательный структурный элемент большинства про-кариотных клеток, который располагается под капсулой или слизистым чехлом или непосредственно контактирует с окружающей средой. На долю клеточной стенки приходится от 5 до 50 % сухого вещества клетки. Это прочная, упругая структура, служащая механическим барьером между протопластом и внешней средой, придающая клеткам определенную, при-сущую им форму и поддерживающая высокое осмотическое давление в клетке. Концентрация солей в клетке, как правило, намного выше, чем в ок-ружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка механически защищает клетку от проникновения в нее избытка воды, то есть сдерживает высокое осмо-тическое давление в клетке. Она участвует в процессе деления клетки и транспорте метаболитов. Клеточная стенка прокариот резко отличается от таковой у эукариот как по строению, так и по химическому составу. Она содержит специфиче-ские полимерные комплексы, которые остутствуют в других структурах клетки. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным признаком при идентификации. зависимости от строения клеточной стенки прокариоты, относя-щиеся к бактериям, делятся на две большие группы. В 1884 г. датский уче-ный Х. Грам предложил метод окраски (впоследствии этот метод стали на-зывать «окраска по Граму»), в результате которого бактерии делятся на грамположительные (сине-фиолетовый) и грамотрицательные (красный цвет). Если фиксированные бактерии окрасить сначала кристаллическим фиолетовым, а затем йодом, то образуется окрашенный комплекс (генциа-новый фиолетовый в комплексе с йодом). В зависимости от строения кле-точной стенки при последующей обработке спиртом этот комплекс либо удерживается, либо вымывается. Если бактерии остаются с сине - фиолетовой окраской, то это свидетельствует о том, что обработка окра-шенного по Граму мазка бактерий спиртом вызывает сужение пор в пепти-догликане и тем самым задерживает краску в клеточной стенке. То есть бактерии окрашиваются грамположительно. 34 Наоборот, грамотрицательные бактерии после воздействия спиртом утрачивают краситель, обесцвечиваются и при обработке фуксином окра-шиваются в красный цвет вследствие меньшего содержания пептидоглика-на (1 - 10 % массы клеточной стенки). В состав клеточной стенки бактерий входят семь различных групп химических веществ (табл. 3). Таблица 3 Химический состав клеточных стенок грамположительных и грамотрицательных бактерий (Rose, 1971 г.; Freer, Salton, 1971 г.)
Примечание. (+) -присутствуют; (-) -отсутствуют; (±)-не у всех. Клеточные стенки грамположительных и грамотрицательных бакте-рий резко различаются по химическому составу и по ультраструктуре. грамположительных бактерий клеточная стенка толще (от 20 до 80 нм), чем у грамотрицательных, и пептидогликан (синонимы муреин, мукопептид) составляет основную массу ее вещества (от 40 до 90 %). Под электронным микроскопом она выглядит как гомогенный электронно-плотный слой. Пептидогликан представлен параллельно расположенными молеку-лами гликана, состоящего из остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью (цв. вклейка, рис. III). Гликановые молекулы связаны поперечной пептидной связью. От-сюда название этого полимера - пептидогликан. Основу пептидной связи составляют тетрапептиды, состоящие из чередующихся L- и D-аминокислот, например L-аланин - D-глутаминовая кислота - мезодиами- 35 нопимелиновая кислота - D-аланин. В пептидогликане грамположитель-ных бактерий вместо мезодиаминопимелиновой кислоты часто содержится L-диаминопимелиновая кислота или лизин. Элементы гликана (ацетилглю-козамин и ацетилмурамовая кислота) и аминокислоты тетрапептида (мезо-диаминопимелиновая и D-глутаминовая кислоты, D-аланин) являются от-личительной особенностью бактерии, поскольку отсутствуют у животных и человека. Пептидогликан ковалентно связан с тейхоевыми кислотами (от греч. teichos - стенка). Это уникальный класс химических соединений, пред-ставляющих собой полимеры, построенные на основе рибита (пятиатомно-го спирта), остатки которого соединены между собой фосфодиэфирными связями. Поскольку это длинные линейные молекулы, они пронизывают весь слой пептидогликана, достигая поверхности клеточной стенки, и яв-ляются основными антигенами грамположительных бактерий. Остающие-ся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона, таким образом, они определяют поверхностный за-ряд клетки. Сахарные компоненты тейхоевых кислот входят в состав ре-цепторов для некоторых бактериофагов и определяют возможность ад-сорбции фага на клеточной поверхности. клеточной стенке грамположительных бактерий содержится не-большое количество полисахаридов, липидов и белков. Входящие в состав клеточной стенки полисахариды, липиды могут ковалентно связываться с ее макромолекулами в отличие от белков, кото-рые формируют на ее внешней поверхности отдельный слой. Таким образом, основными компонентами клеточной стенки грампо-ложительных бактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые, ковалентно связываясь, об-разуют сложную структуру с весьма упорядоченной пространственной ор-ганизацией. Строение клеточной стенки у грамотрицательных бактерий на- много сложнее. У них обнаружена многослойная клеточная стенка. В ее состав входит гораздо большее число макромолекул разного химического типа (рис. 4). Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к цитоплазматической мембране. Для разных видов грамотрицательных бактерий его содержание колеблется в широких пределах и существенно меньше ( 5 - 1 0 %), чем у грамположительных 36 бактерий. Химическая структура пептидогликана грамотрицательных бак-терий в основном сходна со структурой пептидогликана грамположитель-ных бактерий. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки - наружная мембрана. Она состоит из фосфолипи-дов, типичных для элементарных мембран, белков, липопротеина и липо-сахарида. 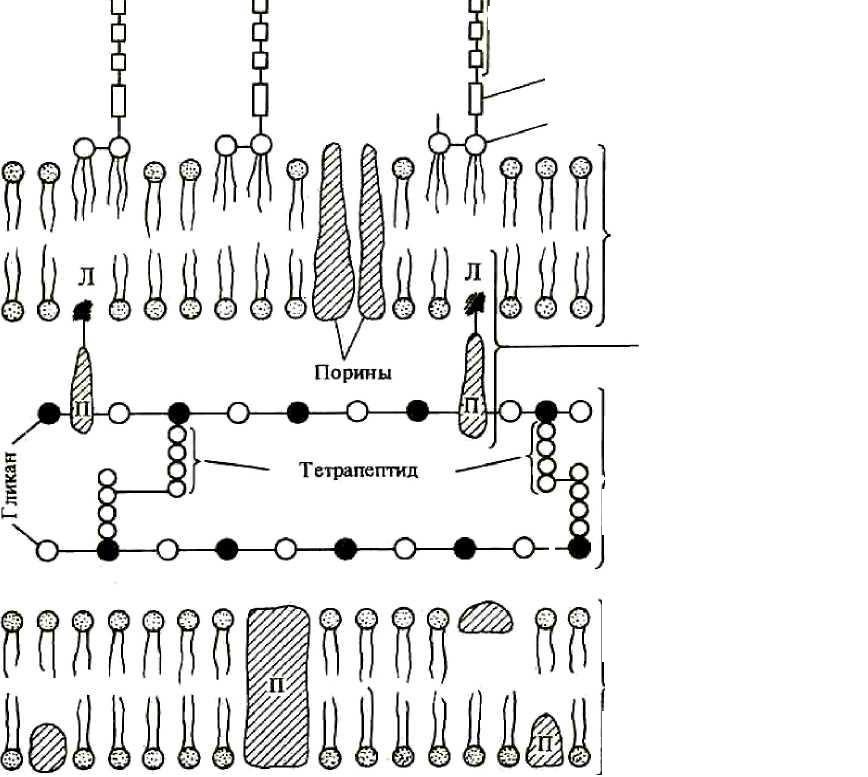 О-Спшифическая цепь ЛПС ЯдроЛПС Л Наружная мембрана Литтопротеин Пегггидогликак иттид А Рис. 4. Схема строения клеточной стенки и Цитотигазматическая мембрана цитоплазматической мембраны грамотрицательных бактерий: Л- липид; П- протеин; ЛПС - липополисахарид Специфическим компонентом наружной мембраны является липопо-лисахарид сложного молекулярного строения, занимающий около 30 - 40 % поверхности и локализованный во внешнем слое. Он состоит из трех компонентов: липидаА, базисной части, или ядра, и О-специфической це-пи полисахарида, образованной повторяющимися идентичными олигоса- 37 харидными последовательностями. Липополисахарид закреплен в наруж-ной мембране липидом А, придающим токсичность липополисахариду, отождествляемому поэтому с эндотоксином. От липидаА отходит базисная часть липополисахарида. Наиболее постоянной частью ядра липополиса-харида является кетодезоксиоктоновая кислота. О-специфическая цепь, от-ходящая от ядра липополисахарида, определяет серогруппу, серовар (раз-новидность бактерий, выявляемая с помощью иммунной сыворотки) выде-ленного штамма бактерий. Таким образом, с понятием липополисахарида связаны представления об О-антигене, по которому можно дифференциро-вать бактерии. Помимо типичной клеточной стенки грамотрицательных бактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной мембраны. Но до настоящего времени не ясно, относятся ли они клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла. Атипичные клеточные стенки прокариот. У некоторых скользя- щих бактерий (миксобактерии, флексибактерии), способных в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, была обнаружена нетипичная для грамотрица-тельных бактерий эластичная клеточная стенка. Объяснением такой гибко-сти клеточной стенки этих бактерий может быть особенность строения пептидогликанового компонента (низкая сшитость этого компонента кле-точной стенки). метанобразующих архей клеточные стенки содержат пептидогли-кан особого химического строения. У других представителей этой группы клеточная стенка состоит исключительно из кислого гетерополисахарида; у некоторых экстремально галофильных, метанобразующих и ацидотер-мофильных архей - только из белка. Археи с клеточной стенкой белковой природы не окрашиваются по Граму, остальные типы архей дают грампо-ложительную реакцию. Прокариоты, не имеющие клеточной стенки. Впервые бактерии, не имеющие клеточной стенки, были обнаружены при воздействии на них лизоцимом - ферментом из группы гликозидаз, содержащимся в яичном белке, слюнной жидкости и выделяемом некоторыми бактериями. 38 Бактерии с частично (сферопласты) или полностью (протопласты) утраченной клеточной стенкой можно получать при воздействии опреде-ленными химическими веществами в лабораторных условиях. При нару-шении синтеза клеточной стенки бактерий под влиянием фермента лизо-цима или пенициллина, а также защитных факторов организма образуются клетки с измененной, часто шаровидной, формой (протопласты). После удаления ингибитора синтеза клеточной стенки такие изме-ненные бактерии могут реверсировать, то есть приобретать полноценную клеточную стенку и восстанавливать исходную форму. Бактерии сферопластного или протопластного типа, утратившие спо-собность к синтезу пептидогликана под влиянием антибиотиков или дру-гих факторов, но сохранившие способность к размножению, называются L-формами. L-формы могут возникать и в результате мутаций. Они пред-ставляют собой осмотически чувствительные шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериаль-ные фильтры. L-формы могут образовывать многие бактерии - возбудите-ли инфекционных болезней. Функции клеточной стенки прокариот многочисленны, разнооб-разны и очень важны для клетки: механическая защита клетки от воздействия окружающей среды; поддержание ее внешней формы; обеспечение возможности существования в гипотонических растворах; транспорт веществ; образование периплазматического пространства у грамотрицатель-ных бактерий, заполненного раствором со специфическими транспортны ми белками и гидролитическими ферментами. У грамположительных бак терий эти ферменты выделяются в окружающую среду, чтобы не происхо дило самопереваривание собственных молекул. ЗНАЧЕНИЕ ПОВЕРХНОСТНЫХ КОМПОНЕНТОВ КЛЕТКИ Все поверхностные компоненты прокариотной клетки имеют боль-шое значение, так как они осуществляют контакт клетки с окружающей средой и позволяют реагировать на внешние воздействия так, чтобы под-держивать собственное существование и выживание в окружающей среде. Их свойства определяются молекулярным составом цитоплазматиче-ской мембраны и клеточной стенки, включая липополисахарид. 39 Наиболее важные функции, которые осуществляют прокариоты, ис-пользуя свои поверхностные компоненты: образуют барьеры проходимости, которые позволяют осуществ лять избирательный проход питательных веществ и исключение вредных веществ; вырабатывают "адгезины", которым свойственно прикрепляться к определенным поверхностям или тканям; выделяют ферменты, чтобы добиться определенных реакций на поверхности клетки, важной для выживания прокариот; выделяют белки, которые могут быть ответом на температуру, ос мотическое давление, соленость, свет, кислород, питательные вещества и т.д., или являться сигналом для генома клетки, который даст правильный ответ на новые воздействия окружающей среды. Область Б - плазматическая мембрана и цитоплазматическая область, которая содержит геном клетки (ДНК) и рибосомы МЕМБРАНЫ Химический состав мембран. Мембраны бактерий структурно по-добны мембранам клетки эукариот, за исключением того, что бактериаль-ные мембраны состоят из насыщенных и мононенасыщенных жирных кислот (редко полиненасыщенных жирных кислот) с 16 - 18 углеродными атомами и обычно не содержат стерины. Содержимое клетки отделяется от клеточной стенки цитоплазмати-ческой мембраной - обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспо-собности. На долю цитоплазматической мембраны приходится 8 - 15 % сухого вещества клетки. У большинства прокариот цитоплазматическая мембрана является единственной мембраной. В клетках фототрофных и ряда хемотрофных прокариот содержатся также мембранные структуры, располагающиеся в цитоплазме и получившие название внутриплазмати-ческих мембран. Их происхождение и функции будут рассмотрены ниже. Цитоплазматическая мембрана - белково-липидный комплекс, кото-рый составляют 50 - 75 % белков, от 15 до 45 % липидов и небольшое ко-личество углеводов. Липиды и белки составляют 95 % и более вещества мембран. Главным липидным компонентом бактериальных мембран явля-ются фосфолипиды - производные 3-фосфоглицерина. Хотя у прокариот 40 найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактери-альных мембранах различные гликолипиды. Стерины отсутствуют у боль-шинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 1 0 - 3 0 % хо-лестерина, поглощаемого из внешней среды, от общего содержания мем-бранных липидов. Из других групп липидов в мембранах обнаружены ка-ротиноиды, хиноны, углеводы. Все липиды бактерий - производные глицерина - содержат один или несколько остатков жирных кислот, состав которых своеобразен. Полине-насыщенные жирные кислоты у бактерий отсутствуют. Исключение со-ставляют цианобактерии, у разных видов которых найдены полиненасы-щенные жирные кислоты типа С16:2, С18:2, С18:3, С18:4. Помимо обычных жирных кислот, то есть обнаруживаемых и в клетках эукариот, в составе мембранных липидов бактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Набор жирных кислот в мембранных липидах также чрезвычайно видоспецифичен. У некоторых грамположительных бактерий С15-жирная кислота с разветвленной цепью может составлять до 90 % всех жирных ки-слот липидов. Главная функция липидов - поддержание механической стабильности мембраны и придание ей гидрофобных свойств. Особый состав липидов обнаружен в мембранах архей. У них не найдены типичные для бактерий эфиры глицерина и жирных кислот, но присутствуют эфиры глицерина и высокомолекулярных С20-, С40-спиртов, а также нейтральные изопреноидные С20-, С30-углеводороды. На долю белков приходится больше половины сухой массы мембран. мембранам с наиболее высоким содержанием белка относятся бактери-альные цитоплазматические мембраны. Цитоплазматическая мембрана E.coli содержит27основных белков и множество минорных белков,но ниодин из основных белков не присутствует в преобладающих количествах. Поскольку цитоплазматическая мембрана прокариот многофункциональна и участвует в осуществлении разнообразных ферментативных процессов, был сделан вывод, что мембранные белки - это, как правило, ферменты. По аминокислотному составу мембранные белки не отличаются от других клеточных белков, за исключением того, что в них содержится мало цис-теина. Некоторые из них являются пермеазами, участвующими в транс-порте веществ. 41 некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. По-видимому, они содержатся не в свободном со-стоянии, а входят в состав гликолипидов и гликопротеинов. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||