Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
Структура мембран. Мембранные липиды всех эукариот и части архей образуют бислои, в которых гидрофильные «головы» молекул обра-щены наружу, а гидрофобные «хвосты» погружены в толщу мембраны. Углеродные цепочки, прилегающие к гидрофильным «головам», довольно жестко фиксированы, а более удаленные части «хвостов» обладают доста-точной гибкостью. У некоторых архей (ряд метаногенов, термоацидофи-лов) мембранные липиды, в состав которых входит С40-спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойная ли-пидная мембрана обладает большей жесткостью по сравнению с бислой-ной. При «биологических» температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят квазикристаллическое состояние. Чем более ненасыщены и разветвлены остатки жирных кислот или чем большее число циклических группировок они содержат, тем ниже температура перехода из жидкостно - кристаллического состояния в квазикристаллическое. «Жидкая» структура мембран обеспечивает определенную свободу молекул белков, что необходимо для осуществления процессов транспорта электронов и веществ через мембрану. Это же свойство обусловливает вы-сокую эластичность мембран: они легко сливаются друг с другом, растяги-ваются и сжимаются. отличие от липидов у мембранных белков нет единого способа структурной организации. 30 - 50 % белка имеет конфигурацию а-спирали, остальная часть находится преимущественно в виде беспорядоч-ного клубка. В зависимости от расположения в мембране и характера связи с липидным слоем мембранные белки условно можно разделить на три группы: интегральные, периферические и поверхностные. Интегральные белки полностью погружены в мембрану, а иногда пронизывают ее на-сквозь. Связь интегральных белков с мембранными липидами очень проч-на и определяется главным образом гидрофобными взаимодействиями. Периферические белки частично погружены в гидрофобную область, а по-верхностные находятся вне еѐ. В первом случае связь с липидами в основ-ном определяется, а во втором - исключительно определяется электроста-тическими взаимодействиями. Помимо этого некоторые белки и липиды в мембране могут быть связаны ковалентно. 42 Одна из функций основных белков - формирование в мембране гид-рофильных диффузионных пор диаметром примерно 1 нм, через которые осуществляется неспецифическая диффузия молекул массой до 600 - 900 Да. То есть через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки выполняют специфические функции: участвуют в облегченной диффузии, активном транспорте веществ и являются специ-фическими рецепторами для фагов. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку желе-зосодержащих соединений. Существует несколько моделей строения мембраны. Наибольшее признание получила модель, учитывающая основные данные, известные о мембранах, согласно которой в липидную основу включены асимметрично расположенные белковые молекулы (см. рис. 4). Некоторые из них обра-зуют скопления на поверхностях липидного би- или монослоя, другие час-тично или полностью погружены в него, третьи пронизывают его насквозь. модели подчеркнута асимметрия строения мембраны, основанная на различиях в химическом строении и расположении молекул белка. При избыточном росте по сравнению с ростом клеточной стенки ци-топлазматическая мембрана образует инвагинаты, то есть впячивания в виде сложно закрученных мембранных структур, называемых мезосомами. Менее сложно закрученные структуры называются внутриплазматически-ми мембранами (ВПМ). Среди ВПМ выделяют несколько видов (табл. 4). Таблица 4 Мембраны прокариот
Примечание. (+) -присутствуют; (-) -отсутствуют; (±)-не у всех. 43 Роль мезосом и внутриплазматических мембран до конца не выясне-на. Полагают, что они участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, а также в секреции веществ, спорообразовании, то есть в процессах, сопровождающихся большой затратой энергии. Функции цитоплазматической мембраны прокариот разнообраз-ны и жизненно важны для клетки: содержит ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ, участвует в синтезе компонентов клеточной стенки; играет важную роль в превращениях клеточной энергии. У бакте рий, источником энергии для которых служат процессы дыхания или фо тосинтеза, в цитоплазматической мембране расположены переносчики це пи электронного транспорта, функционирование которых приводит к гене рированию энергии, используемой затем в клетке по разным каналам, в том числе и для образования энергии в форме АТФ. Содержит значитель ное количество ферментов, системы переноса электронов и является ме стом генерации энергии у бактерий; обеспечивает избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. Осуществляется это с использованием разных механизмов мембранного транспорта. Выделяют 4 типа транспорт ных систем, с участием которых происходит проникновение молекул в бактериальную клетку: пассивную диффузию, облегченную диффузию, ак тивный транспорт и перенос химически модифицированных молекул (транслокация групп); участвует в регуляции роста и клеточного деления; участвует в регуляции процессов репликации и сегрегации хромо сом и плазмид (они связаны с ее рецепторами); с ней связаны жгутики и аппарат регуляции их движения; участвует в компартментализации и стабилизации рибосом; участвует в образовании мезосом (участки инвагинации в цито плазму, они открыты в периплазматическое пространство). ПЕРИПЛАЗМАТИЧЕСКОЕ ПРОСТРАНСТВО Периплазматическое пространство, или периплазма, находится меж-ду цитоплазматической мембраной и внутренним слоем пептидогликана, ширина его у грамположительных бактерий 10 нм. Поры, содержащиеся в 44 клеточной стенке, открываются в периплазматическое пространство. В не-го открыты мезосомы. Оно играет роль во взаимодействии цитоплазмати-ческой мембраны и клеточной стенки, в нем содержатся различные фер-менты (например, фосфатазы), олигосахариды и др. ЦИТОПЛАЗМА Это сложная коллоидная система, в ней нет эндоплазматического ре-тикулума и других органелл, свойственных эукариотам, она неподвижна. Цитоплазма бактерий занимает основной объем клетки и состоит из рас-творимых белков. У некоторых видов есть микротрубочки - рапидосомы, сходные с микротрубочками простейших, и три типа органелл, окружен-ных белковыми мембранами: газовые пузырьки (у водных прокариот - цианобактерий); хлоробиум-везикулы (аппарат фотосинтеза у фотосинте-зирующих); карбоксисомы, которые содержат фермент карбоксидисмута-зу, необходимый для фиксации СО2 в процессе фотосинтеза. цитоплазме располагается ядерный аппарат - генофор (нуклео-плазма), который не отделен мембраной. Кроме хромосомы имеются плаз-миды, иногда целый комплекс. Хромосома и плазмиды связаны со специ-фическими рецепторами цитоплазматической мембраны. В ней располага-ются рибосомы и все компоненты белоксинтезирующей системы. цитоплазме содержатся различные макромолекулы (тРНК, амино-кислоты, нуклеотиды и др.), могут быть мезосомы, которые участвуют в энергетическом обмене, формировании межклеточной перегородки при делении, спорообразовании и др., включения: капельки нейтральных ли-пидов, воска, серы, полисахариды, поли-Р-масляная кислота и полифосфа-ты (волютин). Они накапливаются при избытке питательных веществ в ок-ружающей среде и выполняют роль запасных веществ для питания и энер-гетических потребностей. Зерна волютина выявляются у дифтерийной па-лочки в виде интенсивно окрашивающихся полюсов клетки. Цитоплазматические элементы бактерий неизменно включают про-кариотическую хромосому и рибосомы. Хромосома - обычно одна боль-шая молекула ДНК, замкнутая в кольцо, более или менее свободно лежа-щая в цитоплазме. Прокариоты иногда обладают небольшими внехромо-сомными циклическими ДНК, названными плазмидами. цитоплазме прокариот рибосомы отчетливо видны в форме гранул. Рибосомы бактерий имеют коэффициент седиментации 7 0S (состоят из 5 0S 45 30S субъединиц)в отличие от рибосом,характерных для эукариотиче-ских клеток, у которых коэффициент седиментации 80S (60S и 40S). По-этому некоторые антибиотики, действие которых основано на подавлении синтеза белка путем связывания их с рибосомами бактерий, не оказывают влияния на синтез белка эукариотических клеток. Функция рибосом состоит в участии в процессе трансляции генети-ческой информации с мРНК при синтезе белка. Синтез белка с помощью 70S-рибосомы происходит в митохондриях эукариотических клеток и хло-ропластах, что является главным аргументом в поддержку точки зрения о том, что эти органеллы произошли от прокариот. ГЕНЕТИЧЕСКИЙ АППАРАТ Генетический аппарат, или нуклеоид, является эквивалентом ядра у бактерий. У прокариот это более компактное образование (диаметром око-ло 2 нм), занимающее центральную область в цитоплазме и не отделенное от нее мембраной. ДНК прокариот имеет существенные отличия в структурной органи-зации от эукариотной ДНК: нуклеоид бактерий не имеет ядерной оболоч-ки, ядрышка и основных белков (гистонов). Вся ДНК клетки (и хромосомная, и плазмидная) образует геном клетки. В течение роста клетки и еѐ деления прокариотная хромосома ко-пируется обычным полуконсервативным способом прежде, чем произой-дет еѐ распределение по дочерним клеткам. Однако процессы мейоза и ми-тоза у прокариот отсутствуют. Репликация и сегрегация (разделение) про-кариотной ДНК координируются мембраной, возможно мезосомами. Обычно в бактериальной клетке содержится одна хромосома, но часто в экспоненциально растущей культуре количество ДНК может достигать массы 3, 4, 8 и более хромосом. Нередко в клетках при действии на них определенных факторов (температуры, рН среды, ионизирующего излуче-ния, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках, как правило, обнаруживается по одной копии хромосомы. Нуклеоид выявляется в световом микроскопе после окраски специ-фическими для ДНК методами по Фельгену или Гимзе. На электронных микроскопических фотографиях ультратонких сре-зов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевид-46 ными структурами ДНК. Несмотря на отсутствие ядерной мембраны, нук-леоид довольно четко отграничен от цитоплазмы. Длина молекулы в развернутом виде может достигать более 1 мм, то есть почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей хромосомы бактериальной ДНК не прослеживается никакой закономерности. Однако, если исходить из то-го, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хро-мосом по дочерним клеткам. Было установлено, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую констан-ту седиментации 1300 - 2000S для свободной и 3200 - 7000S для связанной мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20 - 100 независимо суперспирализованных пе-тель. В обеспечении суперспирализованной организации хромосом участ-вуют молекулы РНК. Хромосомы большинства прокариот имеют молеку-лярную массу в пределах 1-10 - 3-10 Да. В группе микоплазм генетиче- ский материал представлен молекулами, имеющими наименьшее для кле-точных организмов количество ДНК 0,4-109 - 0,8-109 Да, а наибольшее со-держание ДНК обнаружено у нитчатых цианобактерий 8-109 Да. Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками - гистонами. В клетках подавляю-щего большинства прокариот не обнаружено гистонов, поэтому нейтрали-зация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg2+ . В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоно-подобные белки, связанные с ДНК. Содержание пар оснований А+Т и Г+Ц молекуле ДНК является постоянным для данного вида организма и слу-жит важным диагностическим признаком. У прокариот молярная доля ГЦ ДНК колеблется в очень широких пределах: от 23 до 75 %. ПОКОЯЩИЕСЯ КЛЕТКИ Некоторые прокариоты наряду с вегетативными формами образуют и покоящиеся формы - цисты и споры. 47 цисты Цисты образуют бактерии рода Azotobacter, группы Спирохеты, рода Myxococcus, рода Rickettsia и др.У большинства миксобактерий образова-ние цист, называемых также миксоспорами, - закономерная стадия их жиз-ненного цикла (рис. 5, а). После окончания стадии активного размножения клетки миксобактерий собираются вместе и образуют структуры наподо-бие плодовых тел, представляющие собой массу слизи, в которую погру-жены клетки, или дифференцированные структуры, поднимающиеся над поверхностью субстрата на простых или разветвленных стебельках (рис. 5, б). 1 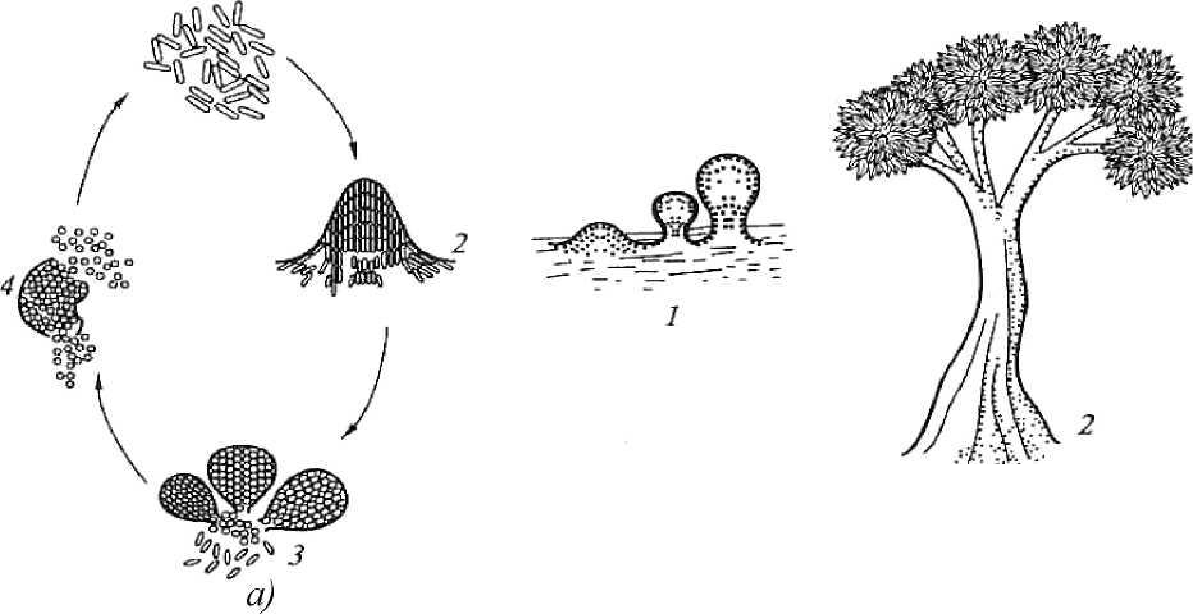 б) Рис. 5. Цикл развития и плодовые тела некоторых миксобактерий: а - цикл развития Myxococcus: 1 - активно размножающиеся вегетативные клетки; 2 - скопление клеток, предшествующее образованию плодового тела; 3 - плодовое тело; 4 - миксоспоры; б - плодовые тела: 1 - Myxococcus; 2 - Choпdromyces (по Schlegel, 1972 г.) [8] Внутри плодовых тел клетки переходят в покоящееся состояние. У одних видов цисты могут морфологически не отличаться от вегетативных клеток, у других их образование сопровождается заметными морфологиче-скими и структурными изменениями: происходит утолщение стенки веге-тативной клетки, в результате чего формируются оптически плотные, бо-лее сильно преломляющие свет, окруженные капсулой укороченные па-лочки или сферические формы (рис. 6, а). Образование миксоспор сопро-вождается синтезом белка, так что сформированная миксоспора содержит 48 около 1/3 заново синтезированного белка. ДНК не синтезируется, а пере-ходит из исходных вегетативных клеток. Генетический аппарат миксоспор может быть представлен тремя или четырьмя копиями хромосомы вегета-тивной клетки. Цисты миксобактерий более устойчивы к нагреванию, высушива-нию, различным физическим воздействиям, чем вегетативные клетки. азотобактера образование цист сопровождается изменением мор-фологии клетки, потерей жгутиков и накоплением в цитоплазме в больших количествах гранул поли-|3-оксимасляной кислоты; одновременно проис-ходит синтез дополнительных клеточных покровов: внешних (экзина) и внутренних (интина) по отношению к клеточной стенке (рис. 6, б), разли- чающихся структурно и химическим составом. 14 15 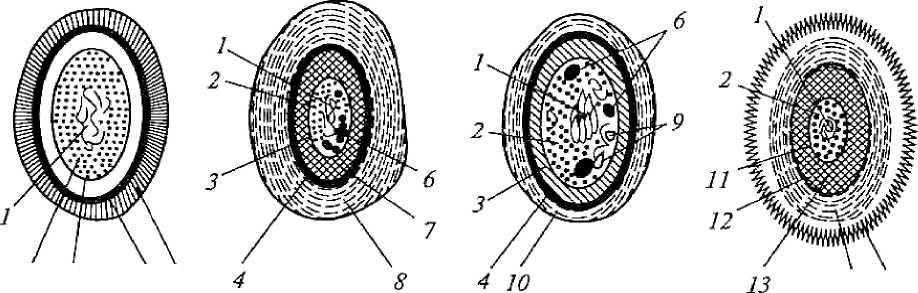 2 3 4 5 а) б) Рис. 6. Строение покоящихся форм прокариот: - миксоспоры миксобактерий; б - цисты азотобактера; в - акинеты циано-бактерий; г - эндоспоры; 1 - нуклеоид; 2 - цитоплазма; 3 - цитоплазматиче-ская мембрана; 4 - клеточная стенка; 5 - капсула; 6 - гранулы запасных ве-ществ; 7 - внутренние покровы (интина); 8 - внешние покровы (экзина); 9 - тилакоиды; 10 чехол; 11 внутренняя мембрана споры; 12 - наружная мембрана споры; 13 - кортекс; 14 - покровы споры, состоящие из нескольких слоев; 15 - экзоспориум (по Дуде, Пронину, 1981 г.) [8] АКИНЕТЫ Покоящимися клетками некоторых цианобактерий, обладающими повышенной устойчивостью к ряду неблагоприятных факторов (высуши-ванию, пониженным температурам), являются акинеты. Они, как правило, заметно крупнее вегетативных клеток, имеют продолговатую или сфериче-скую форму, гранулированное содержимое и толстую оболочку. Образова- 49 ние акинет происходит в период замедления роста и начинается с увеличе-ния клеточных размеров, при этом в цитоплазме накапливаются гранулы запасных веществ (гликогеновых, полифосфатных и особенно крупных цианофициновых), а также карбоксисом. Одновременно происходит утол-щение пептидогликанового слоя клеточной стенки и уплотнение слизисто-го чехла за счет отложения в нем электронно-плотного фибриллярного ма-териала полисахаридной природы (рис. 6, в). Оболочки акинет содержат больше липидов и полисахаридов, а цитоплазма - меньше воды, чем веге-тативные клетки. В цитоплазме при формировании акинет отмечается уве-личение содержания ДНК, рибосом, но уменьшение количества хлорофил-ла и фикобилиновых пигментов. Тилакоиды образуют сложную сетчатую структуру. Скорость фотосинтеза в акинетах ниже, а дыхание выше, чем в вегетативных клетках. Прорастание акинет происходит иногда вскоре по-сле их образования или только после перенесения в свежую питательную среду и может осуществляться двумя путями: иногда в акинете на одном из полюсов формируется пора, через которую выходит проросток, или же прорастание происходит в результате разрыва оболочки акинеты. ЭНДОСПОРЫ Образование эндоспор - процесс, имеющий место только в мире прокариот. Бактериальные эндоспоры - это особый тип покоящихся клеток грамположительных эубактерий, формирующихся эндогенно, то есть внутри цитоплазмы «материнской» клетки (спорангия). Эндоспоры обла-дают специфическими структурами (многослойными белковыми покрова-ми, наружной и внутренней мембранами, кортексом) и устойчивостью к высоким температурам и дозам радиации, летальным в норме для вегета-тивных клеток (рис. 6, г). Эндоспорам свойственно также и особое физиче-ское состояние протопласта. спорообразующим относится большое число бактерий приблизи-тельно из 15 родов, характеризующихся морфологическим и физиологиче-ским разнообразием (табл. 5). Спорообразование, форма и расположение спор в клетке (вегетатив-ной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор может быть овальной, шаровидной; расположе-ние в клетке: терминальное - на конце палочки (возбудитель столбняка), 50 субтерминальное - ближе к концу палочки (возбудитель ботулизма, газо-вой гангрены) и центральное (сибиреязвенная бацилла).
 Примечание: (+) - присутствуют; (-) -отсутствуют; (±)-не у всех. Споры образуются при неблагоприятных условиях существования бактерий, сопровождающихся высушиванием, дефицитом питательных веществ и т.д., при этом внутри одной бактерии образуется одна спора. Поэтому образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие аэробные бакте-рии, у которых размер споры не превышает диаметра клетки, называются бациллами, а спорообразующие анаэробные бактерии,у которых размерспоры превышает диаметр клетки, и они поэтому принимают форму вере-тена, называются клостридиями (от лат. clostridium - веретено). Процесс спорообразования проходит ряд стадий (рис. 7), в течение которых часть цитоплазмы и хромосома отделяются, окружаясь цитоплаз-матической мембраной; образуется проспора. На следующей стадии меж-ду двумя мембранами, покрывающими проспору, формируется толстый слой кортекса (коры). Спорообразование сопровождается интенсивным потреблением проспорой, а затем формирующейся оболочкой споры дипи-колиновой кислоты и ионов кальция. Сформировавшаяся эндоспора состо- 51 ит из протопласта с нуклеоидом, стенки споры, кортекса, оболочки и эк-зоспория. Протопласт споры (ядро) содержит цитоплазматическую мембра-ну, цитоплазму, хромосому, все компоненты белоксинтезирующей и ана-эробной энергообразующей систем. Стенка споры непосредственно окружает ее внутреннюю мембрану представлена пептидогликаном, из которого формируется клеточная стенка прорастающей клетки. 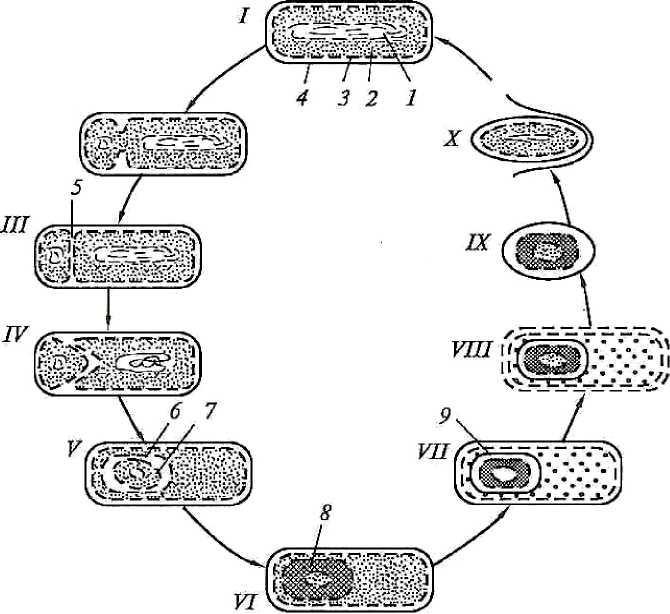 и Рис. 7. Формирование эндоспоры: I - вегетативная клетка; II - инвагинация цитоплазматиче-ской мембраны; III - образование споровой перегородки (септы); IV - формирование двойной мембранной системы образующейся проспоры; V - сформированная проспора; VI- формирование кортекса; VII - формирование покровов споры; VIII - лизис материнской клетки; IX- свободная зре-лая спора; X - прорастание споры; 1 - нуклеоид; 2 - цито-плазма; 3 - цитоплазматическая мембрана; 4 - клеточная стенка; 5 - споровая перегородка; 6 - наружная мембрана споры; 7 - внутренняя мембрана споры; 8 - кортекс; 9 - по-кровы споры (по Дуде, 1974 г.) [8] 52 Кортекс -самый толстый слой оболочки споры.Он состоит из пеп-тидогликана, содержащего мало поперечных сшивок и поэтому очень чув-ствительного к лизоциму. Разрушение кортекса лизоцимом играет пуско-вую роль в процессе прорастания споры. Оболочка споры построена из кератиноподобного белка.Ее плохаяпроницаемость определяет высокую устойчивость спор к действию раз-личных химических веществ. Экзоспорий -липопротеиновая оболочка,содержащая немного уг-леводов. После завершения спорообразования вегетативная часть клетки отмирает, спора высвобождается и длительное время сохраняется в окру-жающей среде до тех пор, пока не возникнут условия, благоприятные для ее прорастания. Специфические элементы споры, включая многослойную оболочку и дипиколинат кальция, обусловливают ее свойства: она долго может сохра-няться в почве, например возбудители сибирской язвы и столбняка - де-сятки лет. В благоприятных условиях они прорастают, проходя три стадии: активацию, инициацию, вырастание. При этом из одной споры образуется одна бактерия. Активация - готовность к прорастанию. Она ускоряется при прогревании при температуре 60 - 80 °С. Инициация прорастания длится несколько минут. Вырастание характеризуется быстрым ростом, сопровождающимся разрушением оболочки споры и выходом проростка. Споры сильно преломляют свет, поэтому хорошо заметны в неокра-шенных препаратах. Генетический контроль спорообразования обеспечивается более чем 40 оперонами, которые представляют дополнительный геном у споро-образующих бактерий. В его составе - до 60 генов. Инициация споруляции связана с геном spoO, мутации в котором делают невозможным образова-ние споры с самых начальных стадий. Транскрипция этого гена запускает последовательную транскрипцию всех оперонов спорового генома. Споро-образующие бактерии обладают механизмами, с помощью которых они распознают определенные изменения в окружающей среде, например уменьшение содержания источников энергии, некоторых аминокислот и др. В ответ в клетке происходят метаболические изменения, которые и за-пускают споруляцию. Одна из особенностей споруляции состоит в том, что на определенном этапе (приблизительно на 3-м часу) происходит синтез небольших кислоторастворимых белков. На их долю приходится около 53 10 - 12 % всех белков споры. В спорах они связываются с ДНК, обеспечи-вая устойчивость к УФ-облучению. В момент прорастания споры эти бел-ки гидролизуются и тем самым снабжают прорастающую спору необходи-мыми аминокислотами. Прорастание споры происходит тогда, когда она получает химиче-ский сигнал. Различные виды спорообразующих бактерий располагают ре-цепторами, распознающими наличие в среде источников энергии, L-аланина, аденозина и др. Связывание с такими эффекторами активирует автолизин (лизоцим), содержащийся в споре, который быстро разрушает пептидогли-кан кортекса. Прорастание спор включает три стадии: Активация является обязательным условием прорастания.Она осу-ществляется различными воздействиями - кислой средой, веществами, со-держащими свободные сульфгидрильные группы, повышением температу-ры, механическим повреждением спор. Начальная стадия. Под влиянием внешних эффекторов происходитактивация автолизина, который разрушает пептидогликан кортекса, в спо-ру поступает вода, спора высвобождается от дипиколината кальция, под воздействием гидролитических ферментов разрушаются другие ее компо-ненты. Стадия роста. После разрушения кортекса и наружных слоев спорыиз нее появляется («выклевывается») растущая новая вегетативная клетка. Она состоит из протопласта споры и ее клеточной стенки. В ней активизи-руются биосинтетические процессы; в результате новая вегетативная клет-ка при наличии необходимых питательных веществ удваивает свою био-массу и делится на две дочерние клетки, которые далее активно размно-жаются, пока этому способствуют условия среды. Процесс прорастания споры контролируется генами как спорового, так и вегетативного генома. Область В - запасные питательные вещества и внутрицитоплазматические включения ВНУТРИЦИТОПЛАЗМАТИЧЕСКИЕ ВКЛЮЧЕНИЯ цитоплазме прокариот обнаруживаются различные включения. Одни из них следует рассматривать как активно функционирующие струк-туры, другие - как продукты клеточного метаболизма, не выделяющиеся 54 наружу, но откладывающиеся внутри клетки. Некоторые цитоплазматиче-ские включения имеют явно приспособительное значение. И наконец, мно-гие из них являются запасными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, потребление наблюдается, когда организм попадает в условия голодания. числу внутрицитоплазматических включений, выполняющих оп-ределенную функцию в фотосинтезе, относят хлоросомы зеленых бактерий и фикобилисомы цианобактерий. В этих структурах локализованы пигмен-ты, поглощающие кванты света и передающие их в реакционные центры, то есть выполняющие роль антенны. Хлоросомы имеют форму продолго-ватых пузырьков длиной 90 - 150 и шириной 25 - 70 нм, окруженных од-нослойной электронно-плотной мембраной толщиной 2 - 3 нм, построен-ной только из белка. Они располагаются в непосредственной близости от ЦПМ, плотно к ней примыкая. В хлоросомах локализованы бактериохло-рофиллы с, d или е. Водорастворимые пигменты белковой природы (фико-билипротеины) цианобактерий содержатся в особых структурах - фикоби-лисомах, расположенных правильными рядами на внешних поверхностях фотосинтетических мембран и под электронным микроскопом имеющих вид гранул диаметром 28 - 55 нм. Карбоксисомы состоят из частиц рибулозодифосфаткарбоксилазы - фермента, катализирующего фиксацию СО2 на рибулозодифосфате в вос-становительном пентозофосфатном цикле. До настоящего времени оконча-тельно не выяснено, в какой форме находится фермент в карбоксисомах: в инертном или функционирующем состоянии. Имеются данные, что в ак-тивно растущей культуре больше фермента находится в растворимой фор-ме. При переходе в стационарную фазу увеличивается доля рибулозоди-фосфаткарбоксилазы в составе карбоксисом. Эти данные указывают на возможную роль карбоксисом как структур, обеспечивающих защиту фер-мента от воздействия внутриклеточными протеазами и его консервирование. внутриплазматическим включениям, имеющим приспособительное значение, относятся магнитосомы, газовые вакуоли, или аэросомы, обна-руженные у водных прокариот. Газовые вакуоли найдены у представите-лей 15 таксономических групп. Это сложноорганизованные структуры, на-поминающие пчелиные соты. Состоят из множества газовых пузырьков, каждый из которых окружен однослойной белковой мембраной и заполнен газом, идентичным газу окружающей среды. Мембраны газовых пузырь-ков проницаемы для газов, но не проницаемы для воды. 55 2.3. Морфология вирусов Вирусы относятся к царству Vira. Это мельчайшие микроорганизмы, не имеющие клеточного строения, белоксинтезирующей системы, содер-жащие только один тип нуклеиновой кислоты (ДНК или РНК). Они отли-чаются особым разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки и затем происходит их сборка в вирусные частицы. Ви-русы, являясь облигатными внутриклеточными паразитами, размножаются цитоплазме или ядре клетки. Сформированная вирусная частица называ- ется вирионом. Морфологию и структуру вирусов изучают с помощью электронного мик-роскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть различной: палочковидной (вирус та-бачной мозаики), пулевидной (вирус бешенства), сферической (вирусы по-лиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги) (рис. 8). 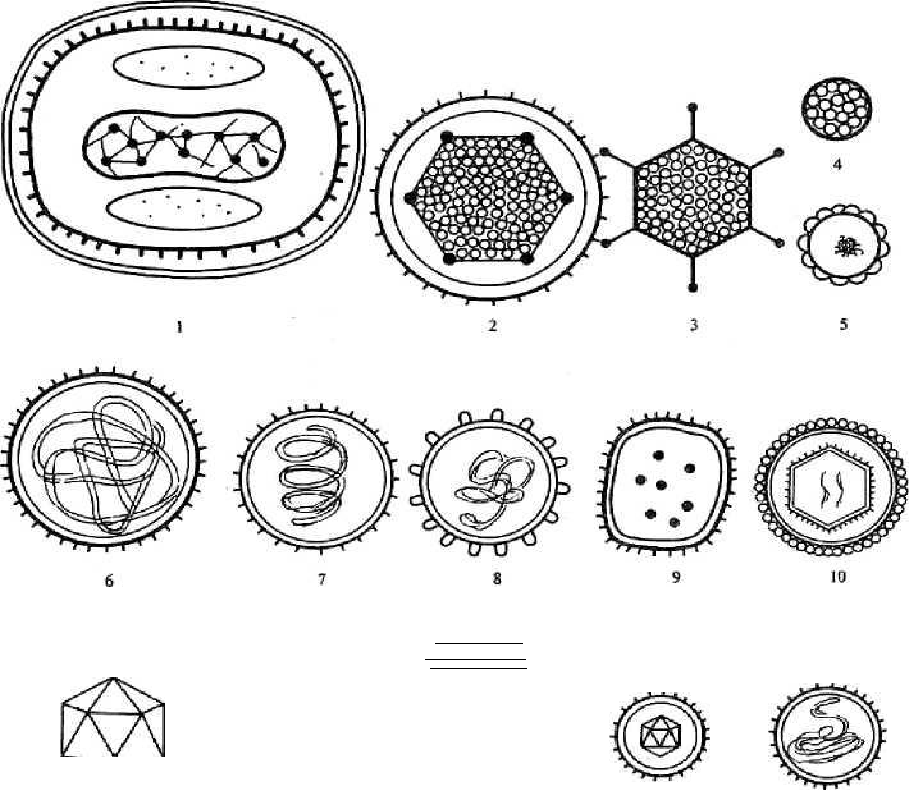 12 Рис. 8. Формы вирионов: - вирус оспы; 2 - вирус герпеса; 3 - аденовирус; 4 - паповавирус; - гепаднавирус; 6 - парамиксовирус; 7 - вирус гриппа; 8 - коронави-рус; 9 - аренавирус; 10 - ретровирус; 11 - реовирус; 12 - пикорнави-рус; 13 - вирус бешенства; 14 - тогавирус, флавовирус; 15 - буньявирус 56 Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одними из самых мелких вирусов яв-ляются вирусы полиомиелита и ящура (около 20 нм), цирковирусы (16 нм), наиболее крупным - вирус натуральной оспы (около 350 нм). Вирусы име-ют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гап-лоидны, то есть имеют один набор генов. Геном вирусов представлен раз-личными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положитель-ным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Име-ются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследствен-ную функцию. Геном вирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (вирусы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плаз-ми ды. Различают просто устроенные (например, вирус полиомиелита) и сложно устроенные (например, вирусы гриппа, кори) вирусы. У просто устроенных вирусов нуклеиновая кислота связана с белковой оболочкой, называемой капсидом (от лат. capsa - футляр). Капсид состоит из повто-ряющихся морфологических субъединиц - капсомеров. Нуклеиновая ки-слота и капсид, взаимодействуя друг с другом, образуют нуклеокапсид. У сложно устроенных вирусов капсид окружен дополнительной липопроте-идной оболочкой - суперкапсидом (производное мембранных структур клетки-хозяина), имеющим «шипы». Капсид и суперкапсид защищают ви-рионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и имму-ногенные свойства вирионов. Внутренние структуры вирусов называют сердцевиной. 57 Для вирионов характерен спиральный, кубический и сложный типы симметрии капсида. Спиральный тип симметрии обусловлен винтообраз-ной структурой нуклеокапсида, кубический - образованием изометриче-ского полого тела из капсида, содержащего вирусную нуклеиновую кислоту. Кроме обычных вирусов, известны и так называемые неканониче-ские вирусы - прионы - белковые инфекционные частицы, имеющие вид фибрилл размером 10-20 х 100 - 200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирус-ной инфекции (болезни Крейтцфельдта - Якоба, куру и др.). Другими не-обычными агентами, близкими к вирусам, являются вироиды - небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||