Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
Рис. 17. Синтез фрагментов Оказаки. Ведущая цепь синтезируется непрерывной, в то время как отстающая цепь синтезируется в виде фрагментов Оказаки Синтез сегмента Оказаки. Вырезание затравочной РНК и замещение ее дезоксирибонуклео-тидами, комплементарными основаниям ДНК-матрицы. Сшивание сегмента Оказаки с предсуществующей нитью ДНК с помощью лигазы. Суперспирализация новых синтезированных участков нитей ДНК. 10. Ревизия ДНК-полимеразой нового синтезированного фрагмента ДНК: нет ли ошибочного включения нуклеотидов. Если получилась ошибка, происходит вырезание ошибочно вклю-ченного нуклеотида с частью этой нити и застройка образовавшейся бреши правильными нуклеотидами. При благоприятных для роста бактерий усло-виях, когда еще не закончился один цикл репликации, могут возникать вторичные и третичные репликативные вилки, благодаря чему в клетке и происходит увеличение массы ДНК и числа копий хромосом. В осуществ-лении процессов репликации ДНК участвует целый комплекс ферментов, образующих единую структуру - реплисому. Генетический контроль реп-ликации ДНК осуществляется большим количеством генов (у Е.соИ их не менее 25), локализованных в самой ДНК; это процесс саморегулируемый. 109 4.3. Формы обмена генетическим материалом у бактерий Помимо основного механизма передачи генов - по наследству (по вертикали), у бактерий существуют следующие формы обмена генетиче-ским материалом по горизонтали, то есть между отдельными особями в популяции клеток: трансформация, трансфекция., трансдукция, конъюгация. Трансформация - перенос генетического материала, заключающий-ся в том, что бактерия-реципиент захватывает (поглощает) из внешней среды фрагменты чужеродной ДНК. Трансформация может быть спонтан-ной или индуцированной. Индуцированная (искусственно получаемая) трансформация происходит при добавлении к культуре бактерий очищен-ной ДНК, полученной из культур тех бактерий, генетические признаки ко-торых стремятся передать исследуемой культуре. Спонтанная трансфор-мация происходит в естественных условиях и проявляется в возникнове-нии рекомбинантов при смешивании генетически различающихся клеток. Она протекает за счет ДНК, попадающей в окружающую среду вследствие лизиса клеток или в результате ее активного выделения жизнеспособными клетками-донорами. процессе трансформации происходят значительные изменения по-верхностных слоев клетки, которые способствуют поглощению ДНК. Ау-толитические ферменты клетки растворяют клеточную стенку в тех участ-ках, где происходит ее синтез. При этом мезосомы через образовавшиеся отверстия соприкасаются с внешней средой, адсорбируют и втягивают внутрь клетки трансформирующую ДНК, где она и вступает в рекомбина-цию с ДНК реципиента. В результате этого образуется мерозигота. Клетка делится, и ее потомки наследуют признаки, полученные от донора и реци-пиента. Однако в других случаях поглощенные фрагменты ДНК разруша-ются нуклеазами клетки-реципиента и трансформации не происходит. Эффективность индуцируемой трансформации во многом зависит от физиологического состояния клеток-реципиентов, а также от размеров трансформирующей ДНК: высокомолекулярная ДНК поглощается труд-нее, чем менее крупные ее фрагменты. Способность к трансформации об-наружена у ряда родов бактерий, но, по-видимому, роль ее в обмене гене-тическим материалом среди бактерий в естественных условиях менее су-щественна, чем роль других механизмов, потому что у многих бактерий имеются особые системы рестрикции и модификации. Эти системы моди-фицируют свою ДНК (чаще всего путем ее метилирования) и разрушают ПО чужеродную ДНК, если она подобным образом не модифицирована. Эф-фективность метода генетической трансформации во много раз повышается том случае, если смесь ДНК и трансформируемых клеток с помощью специального прибора подвергнуть обработке электрическим импульсом. Метод электротрансформации является универсальным, он применим к любым видам бактерий. С помощью этого метода осуществлена транс-формация более 100 видов бактерий. Трансфекция - вариант трансформации бактериальных клеток, ли-шѐнных клеточной стенки, осуществляемый вирусной (фаговой) нуклеи-новой кислотой. С помощью трансфекции удается вызвать у таких бакте-рий вирусную инфекцию. Трансфекцию можно осуществить и с другими (не бактериальными) клетками, если ввести в них чужеродную ДНК, спо-собную рекомбинировать с ДНК этих клеток или способную воспроизво-дить вирионы, или самостоятельно реплицироваться. Трансдукция - перенос генетического материала от клетки-донора к клетке-реципиенту с помощью бактериофагов. Различают трансдукцию неспецифическую и специфическую. Неспецифическая трансдукция - случайный перенос фрагментов ДНК от одной бактериальной клетки к другой. Специфическая трансдукция осуществляется только умеренными фагами, обладающими способностью включаться в строго определенные участки хромосомы бактериальной клетки и трансдуцировать определен-ные гены. Конъюгация. Под конъюгацией понимают процесс обмена генети-ческим материалом (хромосомным и плазмидным), осуществляемый при непосредственном контакте клеток донора и реципиента. Этот процесс контролируется только конъюгативными плазмидами. Аппаратом переноса являются специальные донорные ворсинки (фимбрии, пили), с помощью которых устанавливается контакт между конъюгирующими клетками. Чис-ло донорных пилей у клетки невелико и, очевидно, соответствует числу копий конъюгативной плазмиды в клетке. Донорные ворсинки обнаружи-вают с помощью донорспецифических фагов, которые, адсорбируясь на них, проникают в клетку и вызывают ее лизис. Для каждой группы конью-гативных плазмид существуют свои донорспецифические фаги. Ворсинки выполняют следующие функции: с их помощью устанавливается контакт между донорной и реци-пиентной клетками; 111 они облегчают перенос нити ДНК (она, вероятно, протаскивается через ворсинку); стягивают спаривающиеся клетки, что повышает эффективность конъюгации. Процесс конъюгации протекает через следующие стадии: установле-ние контакта между донором и реципиентом, протаскивание нити ДНК от донора к реципиенту, достройка перенесенной нити ДНК комплементар-ной ей нитью в реципиентной клетке и рекомбинация между переданной хромосомой (ее фрагментами) и хромосомой клетки-реципиента, размно-жение мерозиготы и образование клеток, несущих признаки донора и ре-ципиента. Сущность поверхностного исключения заключается в том, что у бак-терии синтезируются белки наружной мембраны, препятствующие (ис-ключающие возможность) проникновению в клетку, несущую плазмиду, другой, но близкородственной ей плазмиды или подавляющие конъюга-тивную репликацию ее ДНК. Конъюгативная репликация переносимой ни-ти хромосомной или плазмидной ДНК осуществляется также под контро-лем плазмидных генов. Классическим примером конъюгативной плазмиды является половой фактор, или F-плазмида (F - от англ. fertility - плодови-тость). Главная функция этой плазмиды - контроль конъюгации у бакте-рий кишечной группы. Эта плазмида может находиться как в автономном состоянии, так и интегрироваться в хромосому клетки. Находясь в авто-номном состоянии, она контролирует только собственный перенос, при ко-тором клетка, лишенная F-плазмиды, превращается в Р+ -клетку (клетку, содержащую F-плазмиду). F-плазмида может интегрироваться в опреде-ленные участки бактериальной хромосомы, в этом случае она станет кон-тролировать конъюгативный перенос хромосомы клетки. При этом одна из нитей ДНК хромосомы в месте интеграции F-плазмиды разрезается, и ее 5'-конец через донорный мостик начинает протягиваться в клетку-реципиент. Репликация ДНК в этом случае протекает по принципу «кру-тящегося» кольца. Таким образом, конъюгация начинается с установления контакта между донором и реципиентом с помощью донорной ворсинки. Последняя смыкается с рецептором клеточной мембраны клетки - реципиента. Нередко такой контакт устанавливается не только между дву-мя клетками, а между многими клетками, образуя агрегаты спаривания. Поскольку донорный мостик является непрочным, процесс конъюгации может в любой момент прерваться. Поэтому при конъюгации может пере-носиться или часть хромосомы, или, реже, полная хромосома. 112 4.4. Роль генетической информации в регуляции и контроле метаболизма прокариот Цель вегетативной репликации - обеспечить передачу по наследству всех генов. Она происходит последовательно от начала до конца хромосо-мы, в то время как выражение генетической информации, то есть работа генов, подчиняется другой цели, а именно осуществлению за короткий срок жизненного цикла клетки. Поскольку он включает в себя множество биохимических реакций, сопряженных между собой, это предполагает хо-рошо согласованную во времени работу генов. Такая их согласованность возможна лишь при определенном жестком и четком управлении ими. Это управление осуществляется основной структурно-функциональной едини-цей хромосомы - опероном. Гены, участвующие в реализации генетической информации,разде-ляют на структурные, или кодирующие (несущие генетическую инфор-мацию), и регуляторные (осуществляющие управление репликацией того или иного участка ДНК). Оперон -это группа структурных генов-цистронов,физически сцеп-ленных друг с другом и с геном-оператором, который управляет их выра-жением. В состав оперона, как правило, входят структурные цистроны, оп-ределяющие синтез ферментов, которые участвуют в цикле связанных ме-жду собой биохимических реакций. Ген-оператор управляет одновременно всей группой структурныхгенов, которые образуют оперон, иначе говоря, оперон функционирует как самостоятельная единица. В свою очередь, оперон или их группа находит-ся под управлением одного гена-регулятора. Так возникает более сложная структурно-функциональная единица - регулон. Регулон представляет систему,состоящую из гена-регулятора и од-ного или нескольких оперонов, находящихся под контролем одного гена-регулятора. Важным структурным элементом оперона является промотор - об-ласть, с которой взаимодействует РНК-полимераза. составе оперонов могут быть и другие регуляторные элементы: эн-хансеры, аттенуаторы, терминаторы и т. п. Энхансер — генетический элемент,усиливающий транскрипцию опе-рона (от англ. enhance - усиливать). 113 Аттенуатор -генетический элемент,ослабляющий работу оперона(от англ. attenuate - разрежать, разбавлять). Аттенуатор - последователь-ность нуклеотидов, расположенная между промоторным операторным уча-стком оперона и его первым структурным опероном; она кодирует лидер-ную РНК, ее длина около 150 пар нуклеотидов. Терминатор (от англ.terminate -заканчивать)-особый участок вструктуре аттенуатора (лидерной последовательности), от которого зави-сит образование участка мРНК, блокирующего синтез лидерной РНК пе-ред началом первого структурного гена соответствующего оперона. Модель лактозного оперона служит классическим примером орга-низации и работы оперона. Лактоза - дисахарид, она состоит из галактозы глюкозы, соединенных Р-галактозидной связью. Поэтому фермент, раз-рушающий эти связи, получил название Р-галактозидазы. Лактозный оперон (рис. 18) содержит гены, которые контролируют синтез ферментов, участвующих в превращении лактозы: Р-галактозидазу, галактозидпермеазу и тиогалактозидтрансацетилазу. Ген-оператор управ-ляет одновременно выражением всей группы этих генов. В его составе со-держится промотор, с которым взаимодействует РНК-полимераза. 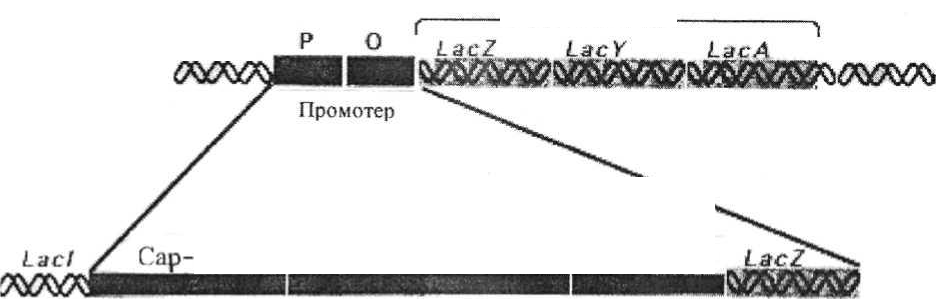 Структурные Регуля горный ген Оперон Р! 1К-полимеразы сайт Сайт взаимодействия Область оператора ас/ Рис. 18. Структура лактозного оперона и элементы его регуляции: I - ген- регулятор; P - промотор; O - оператор; гены, контролирующие синтез: Z - Р-галактозидазы; Y - галактозидпермеазы; A - тиогалактозид-трансацетилазы Работа оперона находится под негативным контролем гена - регулятора, который следит за синтезом белка-репрессора (рис. 19). Бе-лок-репрессор имеет два активных участка: с одним из них взаимодейству- 114 ет индуктор (лактоза), а с помощью другого он прикрепляется к оператору. отсутствие лактозы белок-репрессор связывается с оператором и блоки-рует выражение этого оперона. Когда в среде появляется лактоза, она свя-зывается со вторым активным участком репрессора, это приводит к изме-нению его конформации по типу аллостерического эффекта, и он стано-вится неактивным, репрессия оперона снимается, происходит синтез фер-ментов. Негативный контроль работы лактозного оперона хорошо объясняет сущность феномена индукции: нет индуктора - оперон молчит, его работа заблокирована; появился индуктор - оперон разблокирован и активно ра-ботает.  
 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||