часть_1_2012. Лекции по биологии в 2х кн. Ч. I. Цитология и генетика Под ред проф. Т. В. Викторовой. Уфа, 2012. 192 с., ил
 Скачать 13.87 Mb. Скачать 13.87 Mb.
|

|
Глава 7 История развития генетики. Закономерности наследственности и изменчивости признаков. Основные понятия и термины генетики. законы Менделя. Гипотеза «чистоты» гамет. Дигибридное скрещивание и третий закон Менделя. Виды Генетика изучает закономерности наследственности и изменчивости - два основных свойства живой материи всех организмов.Термин «генетика» был предложен в 1906 г. (Бэтсон). В 1909 г. появились понятия «ген», «генотип», «фенотип» (Иогансен). История развития генетики включает два основных периода: I – доменделевский и менделевский. Доменделевский период
Менделевский период
Условно можно выделить следующие этапы Менделевского периода развития генетики: I этап (1900-1920) – подтверждение законов Менделя на разных объектах. Биохимик Гаррольд писал о распространенности алкаптонурии и изучил химические особенности. Значение: а) продемонстрировал действие законов Менделя (Менделеские болезни, менделирующие признаки). Б) впервые на химическом уровне показал блок гена. 2 этап (1920-1940). Создание хромосомной теории наследственности Морганом и его учениками (Бриджес, Меллер, Стертвант). Заложены основы популяционной генетики (з-н Харди-Вайнберга). 3 этап (1940-1960). Развитие биохимической генетики, молекулярной генетики. 1944 г. Мак-Карти: ДНК – химический субстрат наследственности. Бидл, Татум – «один ген – один фермент» 1953 г. Дж.Уотсон и Ф.Крик – модель ДНК – двунитчатая спираль. Еще раз доказали, что ДНК – субстрат наследственности и изменчивости. Крик, Бреннер – генетический код. 4 этап (1960-1970) – развитие клинической цитогенетики. Денверская и Парижская классификация хромосом. Цитогенетические основы синдрома Дауна и Клайнфельтера). 5 этап (1980-2003) – развитие молекулярной генетики. Совокупная длина ДНК в соматической клетке – 2 метра. 1980 – ПЦР. 1991 – программа «Геном человека». Секвенирование всего генома человека предложено Уотсоном. 6 этап – с 2003 г. – функциональная геномика, протеомика. Основные понятия и термины современной генетики Наследственность – это свойство живых систем передавать из поколения в поколение особенности морфологии, физиологии и индивидуального 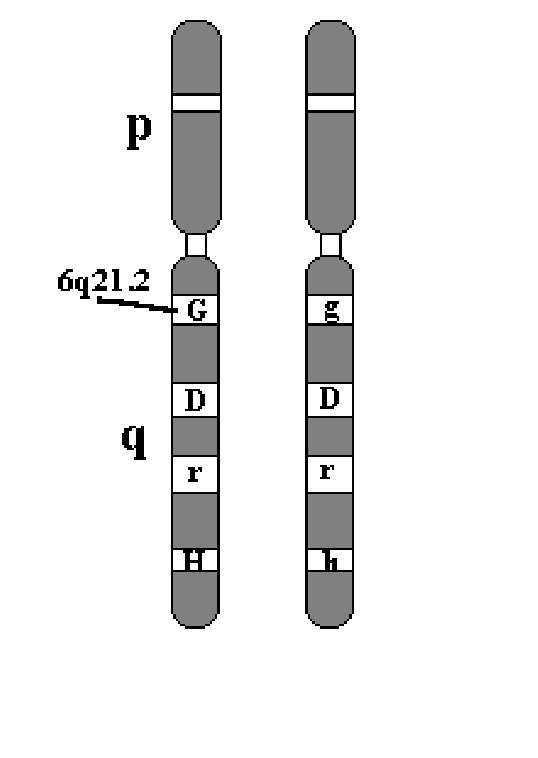 Рис. 7.1. Локализация генов в хромосоме (Аллели: G и g локуса– гетерозиготное состояние по гену, локализованному в локусе 6q21.1: 6 пара гомологичных хромосом, длинное плечо, 2 сегмент, 1 субсегмент ). развития в определенных условиях среды. свойство организмов повторять в ряду поколений сходные признаки. Благодаря наследственности родители и потомки имеют сходство в химическом составе тканей, характере обмена веществ, морфологических признаках и других особенностях. Вследствие этого каждый вид организмов воспроизводит себе подобных из поколения в поколение. Материальными носителями наследственности информации являются гены. Ген - это участок молекулы ДНК, ассоциированный с регуляторными элементами и соответствующий одной единице транскрипции ( один полипептид или один белок). Ген - это функциональная единица наследственности, определяющая развитие какого-либо признака . Геном - совокупность всех генов гаплоидного набора хромосом данного вида особей. •Генотип - совокупность всех генов диплоидного набора хромосом. •Фенотип – внешнее проявление генотипа, реализация генотипа в определенных условиях среды. Гены находятся в хромосомах (рис. 7.1). Гомологичные хромосомы- это хромосомы одинакового размера и морфологии, которые состоят из одних тех же генов, при этом одна из пары гомологичных хромосом является отцовской, другая- материнской. Локус – термин, обозначающий местоположение конкретного гена в хромосоме. Оно постоянно для каждого гена. •Аллель – это варианты одного и того же гена, обусловленные изменениями нуклеотидных последовательностей (м.б. одинаковыми или разными - альтернативными). •Гомозигота - диплоид, содержащий одинаковые аллели данного гена в идентичных локусах гомологичных хромосом (н-р: DD, АА, rr, аа). •Гетерозигота- диплоид, содержащий разные аллели данного гена в идентичных локусах гомологичных хромосом (н-р: Gg, Аа). •Доминантный аллель определяет признак, проявляющийся как в гомо-, так и в гетерозиготном состоянии. • Рецессивный аллельопределяет признак, проявляющийся только в гомозиготном состоянии Альтернативные аллели - разные состояния одного и того же гена (например, аллель А или аллель а). Одинаковые аллели – одинаковые формы гена. Основные закономерности свойств и признаков в поколениях были открыты Г.Менделем в опытах на горохе. Горох – самоопыляемое растение. В своих опытах Мендель использовал гибридологический метод (скрещивал особей с различными генотипами). 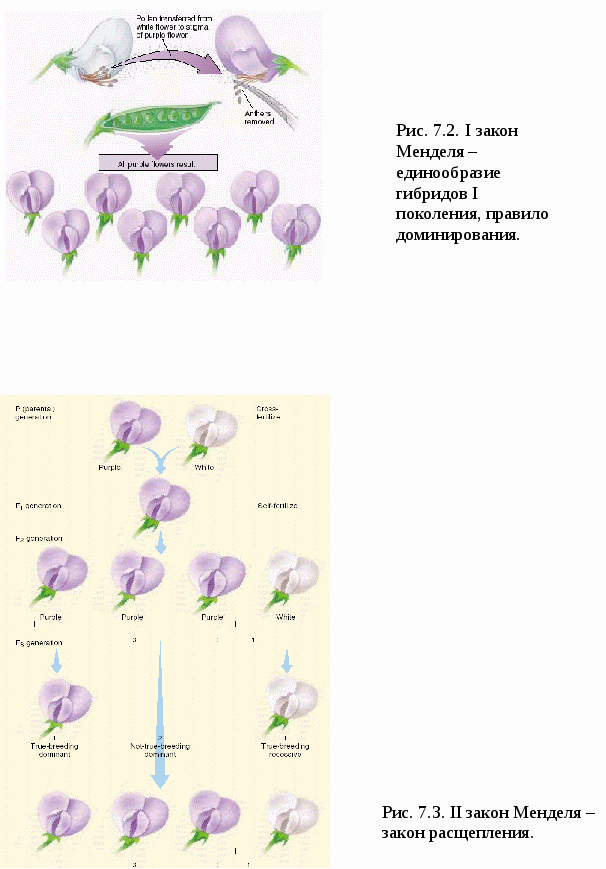 Гибридизация – это скрещивание особей с различными генотипами. Моногибридное скрещивание – скрещивание особей, различающихся по 1 паре альтернативных признаков. Дигибридное - по двум парам. Полигибридное – по многим парам. Особенности гибридологического метода, использованные Менделем.
Чистые линии - это особи, не дающие расщепления по изучаемым признакам, и имеющие только один тип гамет. Примером чистых линий являются особи с генотипами по изучаемым признакам АА; ВВ; ааВВ; ААвв; аавв
I закон Менделя – закон единообразия гибридов I поколения, правило доминирования (рис.7.2). При скрещивании гомозиготных особей, анализируемых по одной паре альтернативных признаков, наблюдается единообразие гибридов I поколения. (Единообразие обусловлено доминированием аллеля А над аллелем а). Затем Мендель скрестил гибридов I поколения между собой. II закон Менделя – закон расщепления (рис.7.3). При скрещивании гибридов I поколения во втором поколении наблюдается расщепление в соотношении 3:1 ( по генотипу 1 : 2 : 1). Особи, содержащие хотя бы один доминантный ген А, имели желтую окраску семян (явление доминирования), а оба рецессивных гена (аа) - зеленую. То есть появились формы, свойственные прародителям. Впоследствии, в 1902 г., после открытия мейоза, Бэтсон для объяснения II закона Менделя предложил цитологическое обоснование и гипотезу «чистоты» гамет: 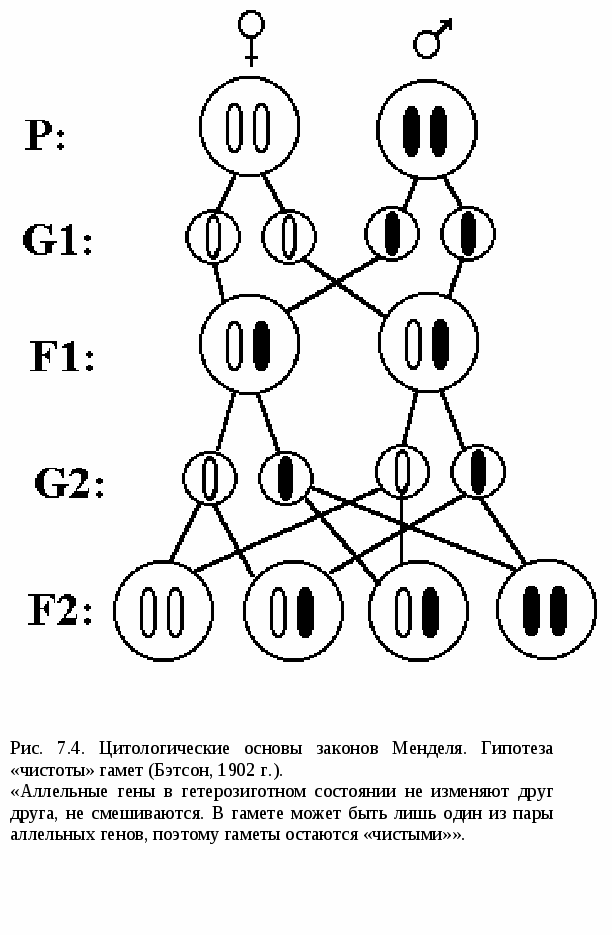 Аллельные гены в гетерозиготном состоянии не изменяют друг друга, не смешиваются. В гамете может быть лишь один из пары аллельных генов, поэтому гаметы остаются «чистыми» (рис.7.4). *Гамета чиста, т.к. в ней находится только одна хромосома. *Вследствие независимого расхождения гомологичных хромосом и хроматид в мейозе из каждой пары аллелей в гамету попадает только один ген. *Аллельные гены находятся в гетерозиготном состоянии. *При оплодотворении гаметы, несущие доминантный и рецессивный признаки, свободно и независимо комбинируются. Для объяснения результатов скрещивания, проведённого Менделем, Бэтсон в 1902 г. предложил, так называемую, гипотезу « чистоты гамет». Изучив наследование 1 пары аллелей, Мендель решил проследить наследование 2-х признаков одновременно. Для этого он использовал гомозиготные растения гороха, различающиеся по 2-м парам альтернативных признаков: цвету (жёлтые и зелёные) и форме (гладкие и морщинистые. В результате в 1 поколении он получил все растения с жёлтыми гладкими семенами, т.е. было показано, что закон единообразия гибридов 1 поколения проявляется и при полигибридном скрещивании. Затем он опять скрестил гибриды 1 поколения между собой. В потомстве оказалось: 9 частей - жёлтые гладкие - генотип А_В_ 3 части - жёлтые морщинистые - генотип А_ вв 3 части - зелёные гладкие -генотип аа В_ 1 часть - зелёные морщинистые - генотип аавв Этот радикал служит основой для всех видов расщепления и взаимодействия аллелей. Его нужно хорошо запомнить. Отсюда вытекает III закон Менделя – закон независимого наследования и комбинирования. (рис.7.5). При скрещивании гомозиготных организмов, анализируемых по двум (или более) парам альтернативных признаков, во 2-ом поколении наблюдается независимое наследование и комбинирование признаков. Мендель обнаружил, что признаки цвета и формы наследуются независимо друг от друга, а именно: В поколении F2: желтых семян - (9+3=12); зеленых (3+1=4); т.е. 12:4=3:1. Такое же распределение наблюдалось по форме семян: гладких - (9+3=12); морщинистых - (3+1=4). 12:4=3:1. Т.е., и по цвету, и по форме соотношение сохраняется таким же, как и при обычном моногибридном скрещивании. Независимое комбинирование признаков проявляется в том, что оба признака могут сочетаться в зиготе независимо друг от друга: 9:3:3:1 – 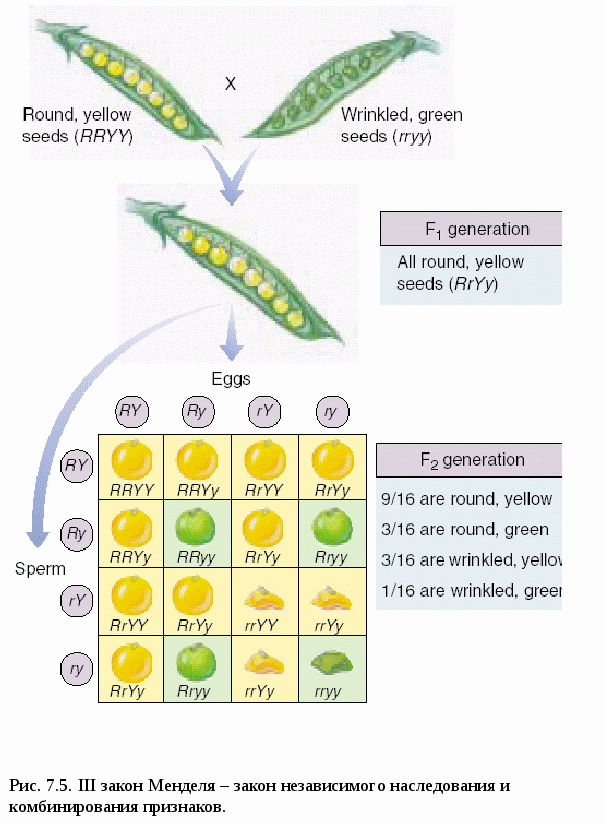 9 – оба признака доминантные 3 – один признак доминантный 3 – другой признак доминантный 1 – оба признака рецессивные Точный количественный учёт признаков позволил Менделю выявить статистические закономерности при полигибридном скрещивании: 1. Количество возможных гамет равно 2n, где n – количество гетерозигот. Например: по 3-м признакам генотип ААВвСс Т.к. n = 2 ( гетерозиготы Вв и Сс ), то количество гамет 22 = 4. При моногибридном скрещивании: АА n= 0 ( гетерозигот нет). 20 = 1 (Один тип гамет А). АаввссДДКК n = 1 Аа n = 1 21 = 2. Получаем два типа гамет (А и а). 2. Число возможных зигот равно 2n1*2n2 , где n1 - число гетерозигот у первого родителя, n2 – у второго. 3. При скрещивании гетерозиготных особей, отличающихся по нескольким параметрам альтернативных признаков, в потомстве наблюдается расщепление по фенотипу (3+1)n, где n – число анализируемых признаков. При дигибридном скрещивании (3+1)2 = 9+3+3+1. На основе своих экспериментов Мендель впервые показал:
Оказалось, что не все признаки подчиняются законам Менделя. Поэтому было предложено понятие менделирующих признаков, т.е. подчиняющихся законам Менделя. У человека более 1000 известных признаков наследуются по законам Менделя (голубые и карие глаза; цвет кожи; веснушки; преобладающая рука; абсолютный музыкальный слух ( дом.+, рец.-); волосы (курчавые волосы доминируют над прямыми волосами); группы крови; моногенные наследственные болезни) и т.д. Анализирующее скрещивание На практике для установления генотипа родительской особи, проявляющей доминантные признаки (которая может быть как гомо-, так и гетерозиготой), её скрещивают с рецессивной формой. Если от такого 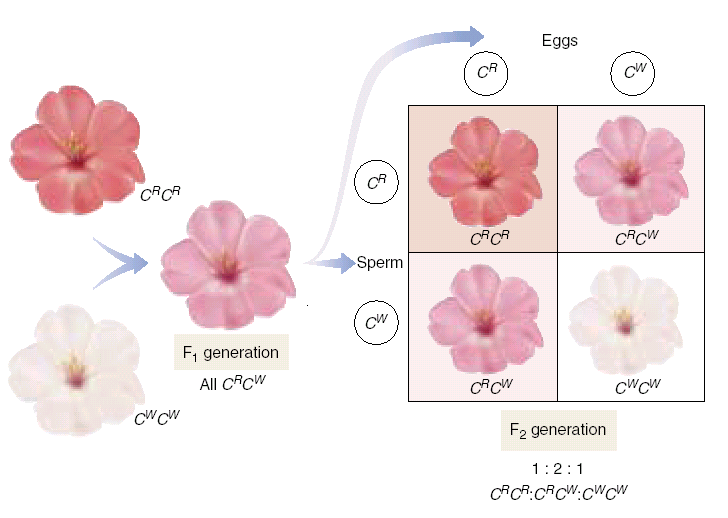 Рис. 7.6.Схема скрещивания при неполном доминировании. 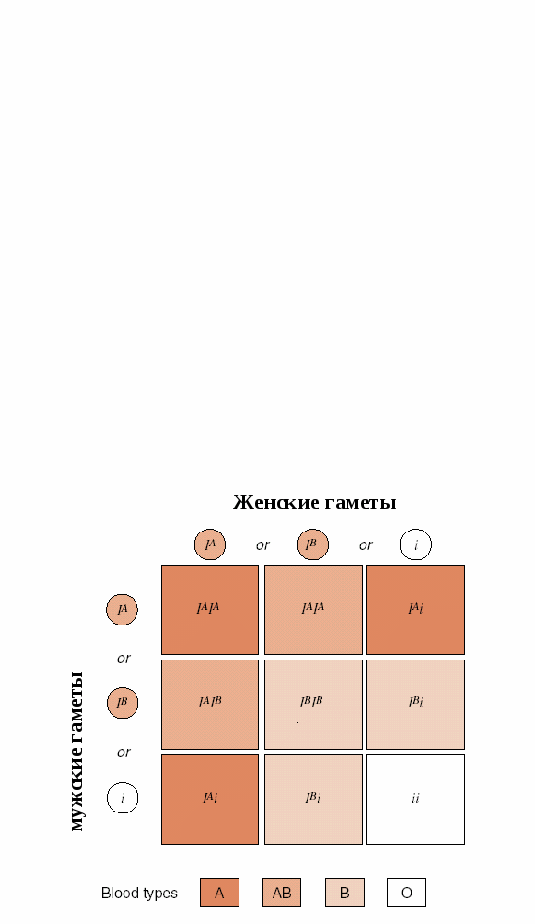 Рис. 7.7. Наследование групп крови системы АВО. скрещивания все потомство окажется однородным, значит анализируемая особь гомозиготна (единообразие гибридов 1 поколения), если произойдет расщепление, то она гетерозиготна. (Пример: мужчина купил чёрного американского дога и захотел выяснить, чистопородный ли он? Если при скрещивании с рецес. геном все чёрные – пес чистопородный (в потомстве Аа), если появится расщепление 1:1 (Аа и аа = 50%:50%) – не чистопородный). Причиной отклонения от законов Менделя являются летальные гены. (АА –чёрные - гибнут, Аа –чёрные, аа – серые). В результате гибели эмбрионов с генотипом АА в родившемся потомстве будет расщепление по фенотипу не 3:1, а 2:1. Но наиболее частой причиной отклонения от законов Менделя является взаимодействие аллелей генов. Различают взаимодействие аллелей одинаковых генов и разных генов. Взаимодействие аллелей одинаковых генов 1. Полное доминирование: А>a – когда доминантный аллель полностью подавляет действие рецессивного аллеля (выполняются законы Менделя). При этом гомозиготы по доминантному признаку и гетерозиготы фенотипически не отличимы (жёлтый горох). 2. Неполное доминирование АА=Аа=аа – домининтный аллель не полностью подавляет рецессивный аллель (рис.7.6). В случае неполного доминирования расщепление по генотипу 1 : 2 : 1 совпадает с расщеплением по фенотипу 1 : 2 : 1 ( красн., розов., белый ).
Доминантный аллель в гетерозиготном состоянии проявляет себя сильнее, чем в гомозиготном. Например : АА – мухи менее плодовиты и живучи, чем Аа. (рецессивная летальная мутация у мух, явление гетерозиса у растений). Множественные аллели Часто в гене может образоваваться несколько разных аллелей. Они возникают вследствие мутаций разных участков в пределах одного гена. Но в генотипе одного диплоидного организма могут находиться только 2 аллеля из серии множественных аллелей ( т.к. гомологичных хромосом две, например: аллель 1 сочетается с аллелем 4, и т.д. 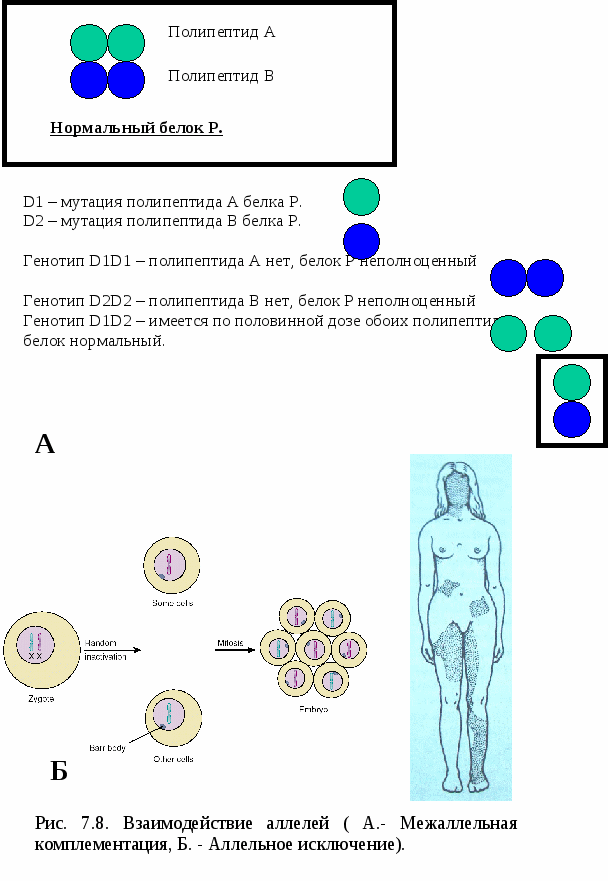 Основные закономерности множественного аллелизма
Примером множественных аллелей у человека могут быть аллели, ответственные за развитие групп крови системы АВО (рис.7.7). Группы крови у человека определяются по наличию на поверхности эритроцитов определённых антигенов. Гены групп крови принято обозначать буквой I. (см. таблицу «Наследование групп крови системы АВО): аллель I0 – на поверхности эритроцитов нет антигенов аллель IА- антиген А аллель IВ –антиген В. При сочетаниях I0I0 – формируется I група крови IАI0 или IАIА – II группа крови IВI0 или IВIВ – III группа IАIВ – IV группа крови На этом примере мы видим явление кодоминирования. 4.Кодоминирование А1+А2=С Оба аллеля (доминантные) равнозначны и в сочетании создают новый признак. Классический пример – 4 группа крови у человека . 5. Межаллельная комплементация (рис.7.8 А) – относится к достаточно редко встречающимся способом взаимодействия аллелей. В этом случае у организма формируется нормальный признак при наличии в генотипе двух мутантых аллелей. Рассмотрим пример: d1d1- мутация полипептида А – неполноценный белок. d2d2 – мутация полипептида В – неполноценный белок. Но! при генотипе d1d2 будут синтезироваться оба полипептида и нормальный белок. 6. Аллельное исключение (рис.7.8 Б). Когда в разных клетках у одной особи проявляются разные аллели. Например, при инактивизации одной из аллелей Х-хромосомы у женщин на некоторых участках кожи отсутствуют потовые железы. Это участки с активной мутантной Х-хромосомой: Х*х - отсутствие потовых желез, х*Х – норма. 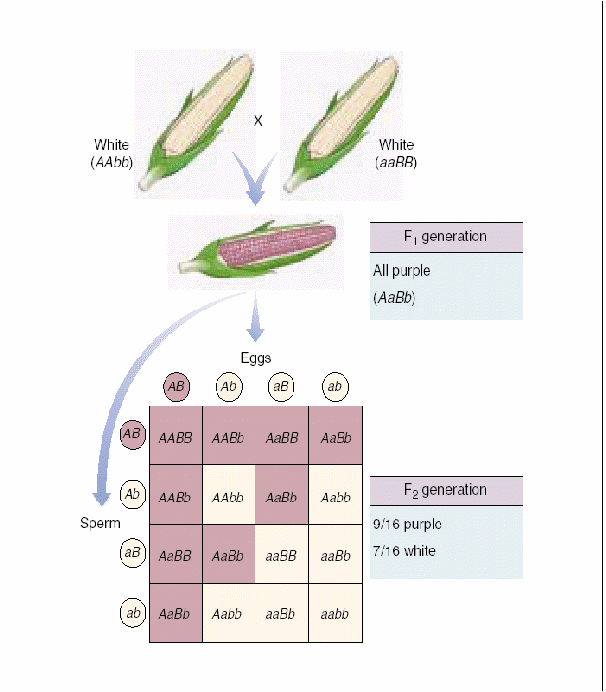 Рис. 7.9. Комплементарное взаимодействие аллелей. Формирование сложных признаков предполагает необходимость взаимодействия аллелей разных генов, занимающих разные локусы (могут быть в разных хромосомах). Виды взаимодействия аллелей разных генов Различают три основных вида такого взаимодействия:
Комплементарность – такой вид взаимодействия доминантных аллелей разных генов, когда признак проявляется при одновременном присутствии в генотипе двух доминантных аллелей) (рис. 7.9.). Чёрный цвет - СС, белый цвет - АА, СА - серые мыши. Душистый горошек – пурпурный цвет А + В У человека: Д – развитие улитки, Е - развитие слух. нерва. Д_ее - глухие ddE_ - глухие ddee - глухие D_E_ - нормальный слух. Эпистаз – это вид взаимодействия аллелей разных генов, при котором один аллель (может быть как доминантным, так и рецессивным) подавляет другой. В этом случае наличие доминантного аллеля А каким-то образом препятствует проявлению доминантного аллеля В, и признак не формируется. Такое взаимодействие аллелей разных генов принято называть эпистатическим. При доминантном эпистазе признак появится только при отсутствии аллеля А, при рецессивном – при отсутствии генотипа аа. Примером эпистатического взаимодействия аллелей может быть подавление у многих видов развития окраски покровов, определяемое одним из аллелей, при наличии в генотипе другого аллеля в доминантном состоянии. Так, у тыквы доминантный аллель В определяет желтую, а рецессивный зеленую окраску. Однако окраска не развивается вообще, если в генотипе имеется эпистатический аллель А. Окраска тыквы: В - жёлтая вв –зелёная А – нет окраски В_аа – жёлтая вв аа – зелёная А_ - нет окраски.  Рис. 7.10. Варианты возможных расщеплений генотипов при различных видах взаимодействия аллелей разных генов. В данном случае доминантный аллель одного гена (репрессор) подавляет действие доминантного аллеля другого гена. Полимерия. Доминантные аллели разных генов влияют на степень проявления одного и того же признака. По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня меланина. В геноме человека имеются четыре разных гена, отвечающие за этот признак. Их обозначают одной буквой (т.к. отвечают за развитие одного признак), но с разными символами, например, ген Р1, ген Р2, ген Р3, ген Р4. Максимальную пигментацию имеют представители негроидной расы, т.к. у них все аллели находятся в доминантном состоянии, формируя генотип Р1Р1 Р2Р2 Р3Р3 Р4Р4. Минимальная пигментация кожи у европеоидов вследствие отсутствия доминантных аллелей (генотип 1р1 р2р2 р3р3 р4р4). Число доминантных аллелей может варьировать от 0 до 8, что обеспечивает разную интенсивность окраски кожи. Полимерное взаимодействие лежит в основе определения количественных признаков (рост, масса, возможно интеллект). Варианты возможных расщеплений генотипов при всех видах взаимодействия неаллельных генов представлены на рис. 7.10. Отсутствие необходимых данных о роли первичных продуктов многих генов в формировании сложных признаков часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образованиях этих признаков. В одних случаях развитие признака при наличии двух аллелей разных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других отсутствие признака, определяемого одним из аллелей при отсутствии другого аллеля в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе. Вероятно, само разделение взаимодействия аллелей на комплементарное и эпистатическое несколько искусственно, ибо во всех этих случаях сложный признак является результатом сочетания в генотипе определенных аллелей соответствующих генов, которые обеспечивают синтез продуктов, участвующих в цепи биохимических преобразований на разных уровнях формирования сложного признака. Плейотропное действие гена – явление, когда один и тот же ген может действовать на различные признаки организма. Например, ген, определяющий рыжий цвет волос, одновременно обусловливает более светлую окраску кожи и появление веснушек. 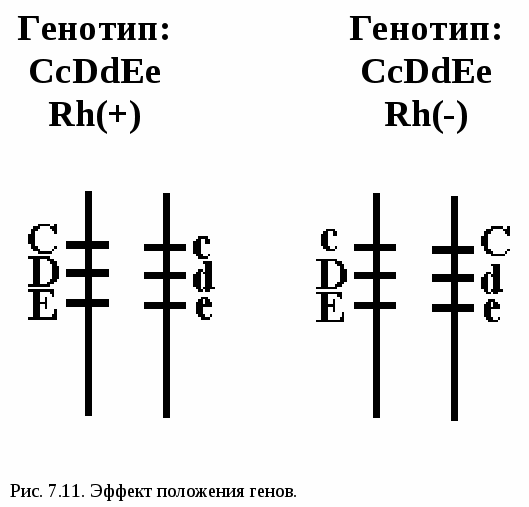 Особый вид представляет взаимодействие, обусловленное местом положения гена в системе генотипа, эффект положения (рис.7.11). Непосредственное окружение, в котором находится ген, может сказываться на характере его экспрессии. Изменение активности гена, наблюдаемое при хромосомных перестройках, нередко связано с перемещением его в другую группу сцепления при транслокациях или изменением его положения в своей хромосоме при инверсиях. Особый случай, очевидно, представляет изменение экспрессии генов в результате деятельности подвижных генетических элементов, активирующих или угнетающих проявление генов, вблизи которых они встраиваются. Наконец, большое значение в объединении генов в единую систему генотипа имеют регуляторные взаимодействия,обеспечивающие регуляцию генной активности. Продукты генов-регуляторов белки-регуляторы обладают способностью узнавать определенные последовательности ДНК, соединяться с ними, обеспечивая, таким образом, транскрибирование информации со структурных генов или препятствуя транскрипции. 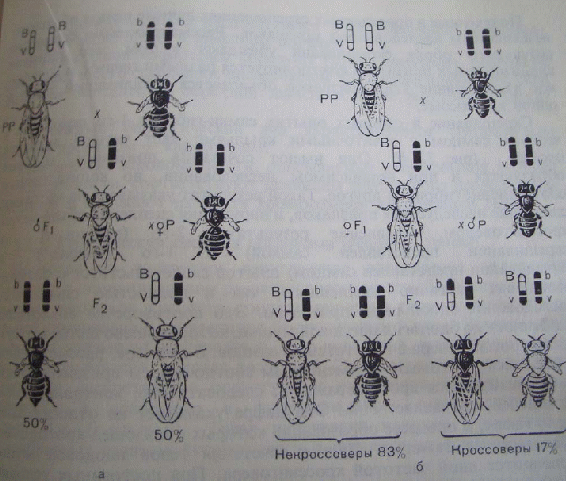 Рис. 8.1. Сцепленное наследование генов окраски тела и состояния крыльев у плодовой мушки (опыты Моргана). |