часть_1_2012. Лекции по биологии в 2х кн. Ч. I. Цитология и генетика Под ред проф. Т. В. Викторовой. Уфа, 2012. 192 с., ил
 Скачать 13.87 Mb. Скачать 13.87 Mb.
|

Регуляция работы генов у эукариотСхема регуляции транскрипции у эукариот разработана Г. П. Георгиевым (1972) (рис. 5.3). Принцип регуляции (обратная связь) сохраняется, но механизмы ее более сложные. Единица транскрипции у эукариот называется транскриптоном. Он состоит из неинформативной (акцепторной) и информативной (структурной) зон. Неинформативная зона начинается с промотора, за которым следуют группа генов-операторов. Информативная зона образована одним структурным геном, разделенным на экзоны (информативные участки) и интроны (неинформативные участки). Заканчивается транскриптон терминатором. Т.о. в состав трантсриптона входят промотор, гены-операторы, 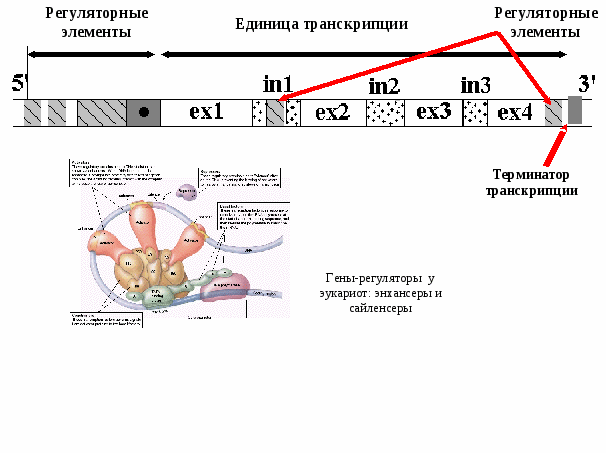 Рис. 5.3. Схема регуляции экспрессии генов у эукариот. Гены эукариот имеют прерывистую структуру (кодирующий фрагмент – экзон, некодирующий фрагмент – интрон) и кодируют только один белок! структурный ген и терминатор. Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для синтеза нескольких белков-репрессоров. Индукторами в клетках эукариот являются сложные молекулы (например, гормоны), для расщепления которых требуется несколько ферментов (многоступенчатые реакции). Когда индукторы освобождают гены-операторы от белков-репрессоров, РНК-полимераза разрывает водородные связи между двумя цепочками ДНК транскриптона и по правилу комплементарности на нем сначала синтезируется большая молекула проинформационной РНК, списывающая информацию как с информативной, так и с неинформативной зон. В дальнейшем в ядре клетки происходит процессинг разрушение неинформативной части РНК. Молекула иРНК формируется посредством сплайсинга (сплавления) отдельных фрагментов ферментами лигазами. Далее иРНК выходит из ядра, идет в рибосомы, где и происходит синтез белка-фермента, необходимого для расщепления индукторов. Включение и выключение транскриптона происходит принципиально так же, как и оперона. Таким образом, по химической организации наследственного материала эукариот и прокариот клетки принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является генетический код. Принципиально одинаковым образом у про и эукариот осуществляется использование наследственной информации, хранящейся в ДНК. Реализация генетической информации Биосинтез белка Одним из центральных процессов метаболизма клетки является синтез белка, т.е. формирование сложной молекулы полипептида из мономеров – аминокислот. Синтезированные в клетке полипептиды либо сами являются полноценными белками, либо входят в состав сложных белков в виде субъединиц. Информация о структуре белков (полипептидов) зашифрована в молекуле ДНК в виде генетического кода (вспомним свойства генетического кода: триплетность, универсальность, специфичность, вырожденность, непрерываемость и неперекрываемость). Как осуществляется перенос этой информации? Какова связь между геном и признаком? Как происходит биосинтез белка? Весь процесс биосинтеза белка можно представить в виде очень простой схемы, которую необходимо хорошо запомнить. Представление о том, что генетическая информация хранится в клетке в виде молекулы ДНК и реализуется благодаря транскрипции в РНК и последующей трансляции в 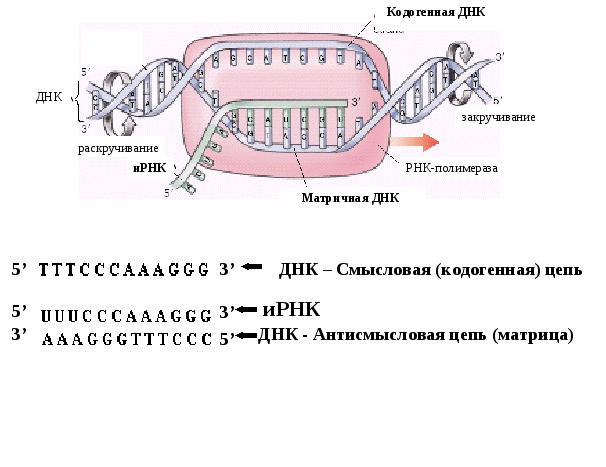 Рис. 5.4.Схема синтеза матричной РНК (транскрипция). Матрицей для транскрипции мРНК служит только одна – антисмысловая (матричная) цепь ДНК, ориентированная в направлении 3’- 5’. Рис. 5.4.Схема синтеза матричной РНК (транскрипция). Матрицей для транскрипции мРНК служит только одна – антисмысловая (матричная) цепь ДНК, ориентированная в направлении 3’- 5’.белок известно, как «Центральная догма молекулярной биологии», которую можно представить в виде простой формулы: ДНКРНК белок. транскрипция трансляция Как видно, функционирование (экспрессия) генов от ДНК до белка реализуется благодаря двум глобальным молекулярно-генетическим механизмам: транскрипции и трансляции. Итак, генная информация у всех клеток закодирована в виде последовательности нуклеотидов в ДНК. Первый этап реализации этой информации состоит в образовании РНК по подобию ДНК, который называется транскрипцией. I этап биосинтеза белка – транскрипция Транскрипция (рис. 5.4) начинается с обнаружения особого участка гена в молекуле ДНК, который указывает место начала транскрипции - промотора с помощью специального фермента РНКполимеразы. После присоединения к промотору РНКполимераза раскручивает прилежащий виток спирали ДНК. Две цепи расходятся и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением правила комплементарности нуклеотидов. В связи с тем, что РНКполимераза способна собирать полинуклеотид только в одном направлении, а именно от 5’ к 3’-концу, матрицей может служить только та цепь ДНК, которая обращена к ферменту своим 3’-концом. Такую цепь называют матричной или антисмысловой. Другая, антипараллельная цепь ДНК, называется кодогенной или смысловой, т.к. последовательность нуклеотидов этой цепи полностью соответствует последовательности РНК и читается в том же направлении, т.е. от 5’ к 3’-концу. Поэтому генетический код иногда пишут по молекуле РНК, иногда – по кодогенной ДНК. Продвигаясь вдоль цепи ДНК, РНКполимераза осуществляет последовательное точное переписывание информации до тех пор, пока она не встречает на своем пути STOPкодонтерминатор транскрипции. У человека три STOP-кодона – TAG, TGA, ТAA (или UAG, UGA, UAA). П этап биосинтеза белка трансляция Трансляция (рис.5.5) включает 3 фазы: инициация, элонгация и терминация. 1 фаза - Инициация фаза начала синтеза полипептида
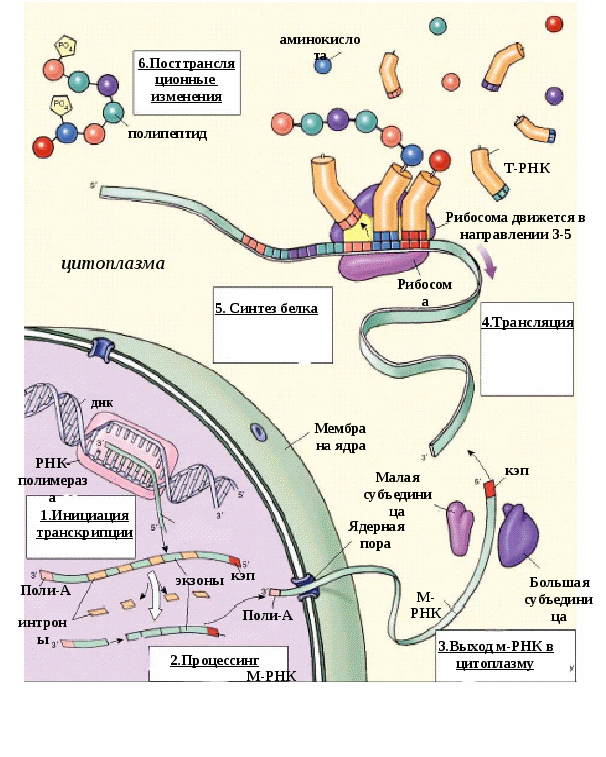 Рис. 5.5.Обобщенная схема процесса экспрессии генетической информации в эукариотической клетке. Рассмотрим, как же проходят в клетке эти процессы. 1) В молекуле любой мРНК вблизи 5’-конца имеется участок, комплементарный последовательности нуклеотидов рРНК малой субчастицы рибосомы. Рядом с этим участком расположен стартовый кодон АУГ, кодирующий аминокислоту - метионин. Малая субчастица рибосомы соединяется с мРНК. Затем происходит объединение малой субчастицы с большой субчастицей, формируется рибосома. В рибосоме образуются два важных участка – пептидильный центр - П-участок и аминоацильный центр – А-участок. К концу фазы инициации П-участок занят аминоацил тРНК, связанной со стартовой аминокислотой - метионином, а А-участок готов принять следующий за стартовым кодон. 2) В рибосомы транспортируются молекулы тРНК. Молекулы тРНК состоят из 7595 нуклеотидов и по форме напоминают лист клена. В своем составе они имеют два активных центра: 1) акцепторный конец, к которому присоединяется транспортируемая аминокислота путем ковалентной связи с затратой энергии 1 АТФ. Формируется аминоацил тРНК. 2) антикодоновая петля, комплементарная кодону мРНК. 2я фаза элонгация удлинение полипептида. Внутри большой субчастицы рибосомы одновременно находятся около 30 нуклеотидов мРНК и только 2 информативных триплета-кодона: один в аминоацильном А-участке, другой в пептидильном П-участке. Молекула тРНК с аминокислотой вначале подходит к Ацентру рибосомы. В том случае, если антикодон тРНК комплементарен кодону мРНК, происходит временное присоединение аминоацил-тРНК к кодону мРНК. После этого рибосома передвигается на 1 кодон по мРНК, а тРНК с аминокислотой перемещается в Пучасток. К освободившемуся Аучастку приходит новая аминоацил-тРНК с аминокислотой и вновь останавливается там в том случае, если антикодон тРНК комплементарен кодону мРНК. Между аминокислотой и полипептидом образуется пептидная связь и одновременно разрушается связь между аминокислотой и ее тРНК, а также между тРНК и мРНК. Освободившаяся от аминокислоты тРНК выходит из рибосомы в цитоплазму. Она готова соединиться со следующей аминокислотой. Рибосома снова перемещается на 1 триплет. 3 фаза терминации завершение синтеза полипептида. Когда на рибосоме появляется один из бессмысленных STOP-кодонов, синтез белка прекращается. При этом к последней аминокислоте присоединяется вода и ее карбоксильный конец отделяется от тРНК. В результате пептидная цепь теряет связь с рибосомой, которая распадается на 2 субчастицы. Глава 6. Мейоз как процесс формирования гаплоидных гамет. гаметогенез. Размножение организмов как механизм, обеспечивающий смену поколений. Основные способы размножения Мейоз Мейоз – это вид деления клеток, при котором из одной диплоидной клетки образуются 4 гаплоидные (рис. 6.1.). Мейоз включает два деления: 1 - редукционное и 2 - эквационное (уравнительное). Мейоз состоит из ряда фаз. Предшествует мейозу интерфаза, в процессе которой происходит синтез ДНК— редупликация хромосом. Редукционное деление начинается с профазы I, которая принципиально отличается от профазы митоза. Профаза I состоит из ряда стадий: лептотена, зиготена, пахитена, диплотена, диакинез. В стадии лептотены (тонких нитей) уже хорошо видны отдельные нити очень тонких (слабо спирализованных) и длинных (в 2—5 раз длиннее метафазных) хромосом. Хромосомы в это время состоят из двух хроматид, соединенных общим участком — центромерой. Это говорит о том, что удвоение хромосом, их редупликация, в основном произошла в интерфазе, предшествующей мейозу. На стадии зиготены (стадия конъюгирующих нитей) хромосомы, одинаковые по размеру и морфологии, т. е. гомологичные, притягиваются друг к другу— конъюгируют. Они соединяются друг с другом наподобие застежки «молния». Такое объединение хромосом-гомологов осуществляется благодаря присущей только мейозу уникальной структуре – синаптонемальному комплексу. Синаптонемальный комплекс обеспечивает тесный контакт между гомологичными сегментами хроматид. Это важное генетическое событие, поскольку конъюгация делает возможным обмен участками между несестринскими хроматидами гомологичных хромосом, приводя к качественному изменению внутренней генетической структуры хромосом. Этот обмен участками между хроматидами гомологичных хромосом получил название «кроссинговер». Каждая пара конъюгирующих гомологичных хромосом образует бивалент. Бивалент, таким образом, состоит из четырех хроматид, поэтому бивалент называют тетрадой. В следующей стадии—пахитене (стадия толстых нитей)—происходит утолщение и укорочение хромосом, главным образом за счет спирализации и конденсации. Хроматиды одной хромосомы—сестринские - хорошо видны. В этой стадии хорошо различима продольная дифференциация хромосом, так что можно идентифицировать каждую хромосому. В следующей стадии—диплотене (стадия двойных нитей) гомологичные хромосомы начинают отталкиваться. Процесс отталкивания начинается в области центромеры и распространяется к концам. В это время Рис. 6.1. Схема мейотического деления клетки. 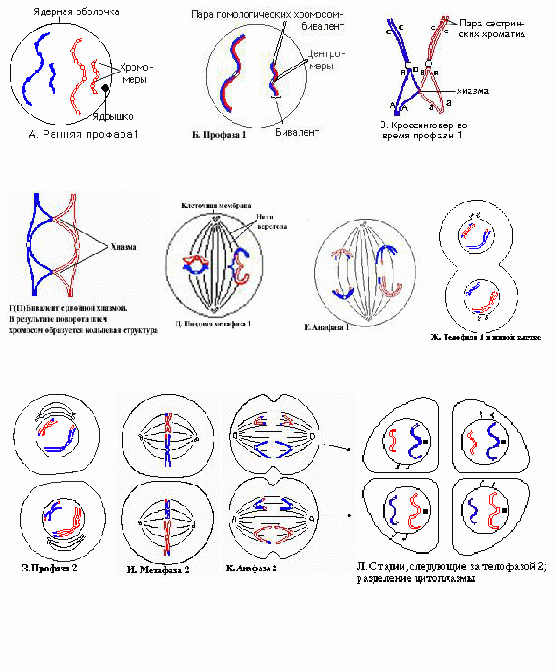 хорошо видно, что бивалент состоит из двух хромосом (откуда и название стадии «двойные нити»). В биваленте обособлены четыре хроматиды, поэтому бивалент называют тетрадой. В это же время становится видно, что тела двух гомологичных хромосом переплетаются. Фигуры перекрещенных хромосом напоминают греческую букву «хи» (χ), поэтому места перекреста назвали хиазмами. Наличие хиазм связано с произошедшим кроссинговером. По мере прохождения этой стадии хромосомы как бы раскручиваются, происходит перемещение хиазм от центра к концам хромосом (терминализация хиазм). Это обеспечивает возможность движения хромосом к полюсам в анафазе. На стадии диакинеза (стадия движения вдаль или стадия расхождения нитей) биваленты, которые заполняли весь объем ядра, начинают перемещаться ближе к ядерной оболочке. К концу диакинеза контакт между хроматидами сохраняется на одном или обоих концах. Исчезновение оболочки ядра и ядрышек, а также окончательное формирование веретена завершают профазу I. В метафазе I биваленты прикреплены в области центромер к нитям веретена, причем центромеры гомологичных хромосом располагаются на противоположных сторонах экваториальной плоскости. Гомологичные хромосомы связаны друг с другом переместившимися к концам хромосом хиазмами. Число бивалентов вдвое меньше, чем число хромосом в соматической клетке организма, т. е. равно гаплоидному числу. В анафазе I к полюсам отходят гомологичные хромосомы из каждого бивалента, т. е. число хромосом у каждого полюса уменьшается вдвое. Центромеры не делятся. В этот редуцированный гаплоидный набор попадает обязательно по одной гомологичной хромосоме из каждого бивалента. Происходит независимая комбинация гомологичных хромосом (отцовские b материнские). В этом принципиальное отличие мейоза от митоза. Телофаза I очень короткая, в процессе ее идет формирование новых ядер. Хромосомы деконденсируются и деспирализуются. Так заканчивается редукционное деление. Цитокинез у многих организмов происходит не сразу после деления ядер, так что в одной клетке лежат два ядра более мелких, чем исходное. Затем наступает стадия интеркинеза, которая отличается от интерфазы тем, что в ней не происходит синтеза ДНК и дупликации хромосом. Интеркинез – это промежуточная стадия между редукционным и эквационным делениями мейоза. Вслед за интеркинезом наступает второе деление мейоза—эквационное. Оно проходит по типу митоза. Эквационное деление состоит из тех же фаз, что и митоз: профаза II, метафаза II (при рассмотрении пластинки с полюса хорошо видно, что число хромосом в клетке гаплоидное, а каждая хромосома состоит из двух хроматид), анафаза II (хроматиды расходятся к полюсам), телофаза II (в каждом ядре—гаплоидное число хромосом, а число ядер в клетке равно четырем). В клетке происходит цитокинез, в результате которого образуются четыре клетки. Итак, в мейоз I вступает диплоидная клетка с удвоенным набором хромосом. В результате мейоза I образуются две гаплоидные клетки с удвоенными хромосомами. В результате мейоза II образуются четыре гаплоидные, генетически разнородные клетки с одинарными хромосомами. Отличия мейоза от митоза (рис.6.2)
Биологическое значение мейоза Мейоз – это механизм, с помощью которого поддерживается постоянство числа хромосом (кариотип) у животных при половом размножении от одного организма к другому. Значение мейоза состоит также в том, что в анафазе I деления мейоза каждая пара хромосом ведет себя независимо от другой пары, в результате чего образуется большое число различных новых комбинаций негомологичных хромосом, т.е. происходит рекомбинация генов на уровне целых хромосом. Благодаря такому механизму достигается большое число новых сочетаний наследственной информации. Оно может быть выражено формулой 2n, где n – число хромосом в гаплоидном наборе. Например, у дрозофилы n = 4 и количество типов гамет, обеспечиваемое комбинацией родительских хромосом в них, равно 24 = 16. У человека n = 23, и разнообразие гамет, обусловленное этим механизмом, соответствует 223, или 8 388 608. Далее в процессе кроссинговера происходит рекомбинация генетического материала, что создает мощный дополнительный резерв наследственной изменчивости. Мейоз 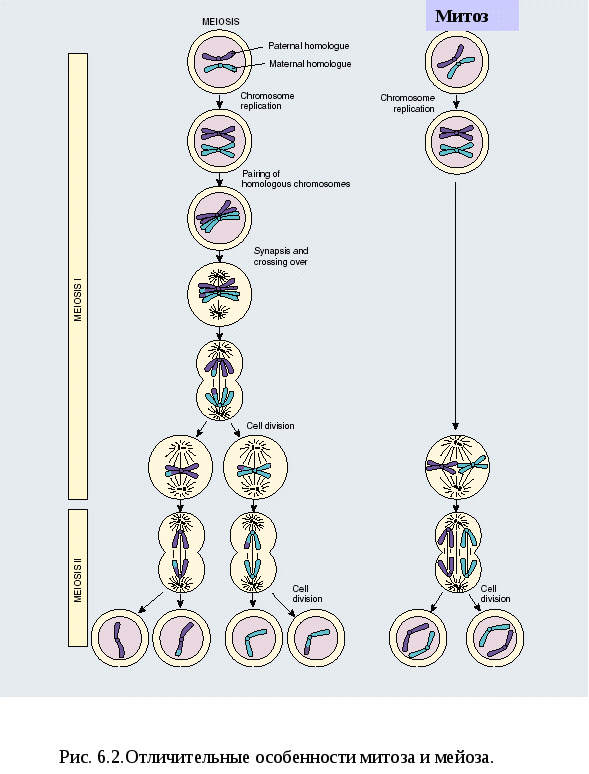 Мейоз лежит в основе образования половых клеток при половом размножении многоклеточных организмов. Развитие гамет происходит в половых железах – гонадах. Формирование мужских половых клеток (сперматозоидов) происходит в семенниках и называется сперматогенезом. Процесс образования женских половых клеток (яйцеклеток) происходит в яичниках и называется овогенезом. Сперматогенез Сперматогенез происходит в мужских половых железах – семенниках (рис. 6.3). 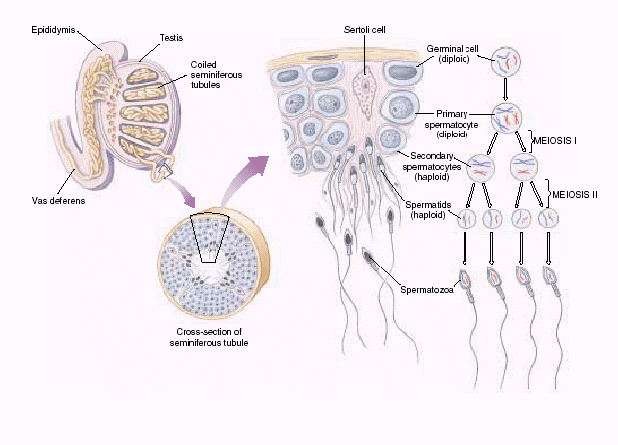 Рис. 6.3. Схема сперматогенеза. Семенник состоит из многочисленных канальцев. На поперечном разрезе через каналец видно, что его стенка состоит из нескольких слоев клеток, которые представляют собой последовательные стадии развития сперматозоида. Наружный слой – сперматогонии. Сперматогонии развиваются их первичных половых клеток, мигрирующих в семенники на ранних стадиях эмбрионального развития. Когда животное достигает периода половой зрелости сперматогонии начинают быстро делиться путем митоза. Это период размножения (I) сперматогониев, который продолжается на протяжении всего периода половой зрелости мужской особи. Часть сперматогониев перемещается в следующую зону – зону роста, расположенную ближе к просвету семенного канальца. В период роста (II) происходит увеличение объема цитоплазмы и клеточных размеров. Важным событием этого периода является репликация ДНК. Хромосомы приобретают двунитчатую структуру (набор хромосом 2n4c) – сперматоциты I порядка. Следующий период – период созревания (III). Основным событием этого периода является мейоз, включающих два последовательных деления (редукционное и эквационное). В результате редукционного деления образуются сперматоциты II порядка (набор n2c), затем – сперматиды (хромосомный набор nc). Сперматиды – клетки с гаплоидным набором хромосом. Процесс сперматогенеза завершается периодом формирования (IV). Сперматиды перемещаются ближе к просвету семенного канальца, меняется их форма, формируются зрелые сперматозоиды – клетки, способные к передвижению, которые выходят в просвет канальца. Т.о. сперматогенез включает 4 периода: размножение, рост, созревание и формирование. Из одного сперматогония образуется 4 зрелых сперматозоида. Морфология сперматозоидов.Сперматозоиды – очень маленькие клетки, обладающие способностью к передвижению. Сперматозоид имеет головку, шейку и хвост. На переднем конце головки расположена акросома, состоящего из видоизмененного комплекса Гольджи. В ней содержится фермент для растворения оболочка яйца. Основную массу головки занимает ядро. В шейке находятся центриоль и много митохондрий. От шейки отрастает хвост, представляющий собой жгутик – специализированный органоид для передвижения. Овогенез Процесс развития женских половых клеток (яйцеклеток) называется овогенезом. Овогенез происходит в женских половых органах – яичниках (рис. 6.4). Периоды развития яйцеклеток сопоставимы с Периодами сперматогенеза. Овогонии развиваются из первичных половых клеток, мигрирующих в яичник на ранней стадии эмбриогенеза. Период размножения (I) овогоний у 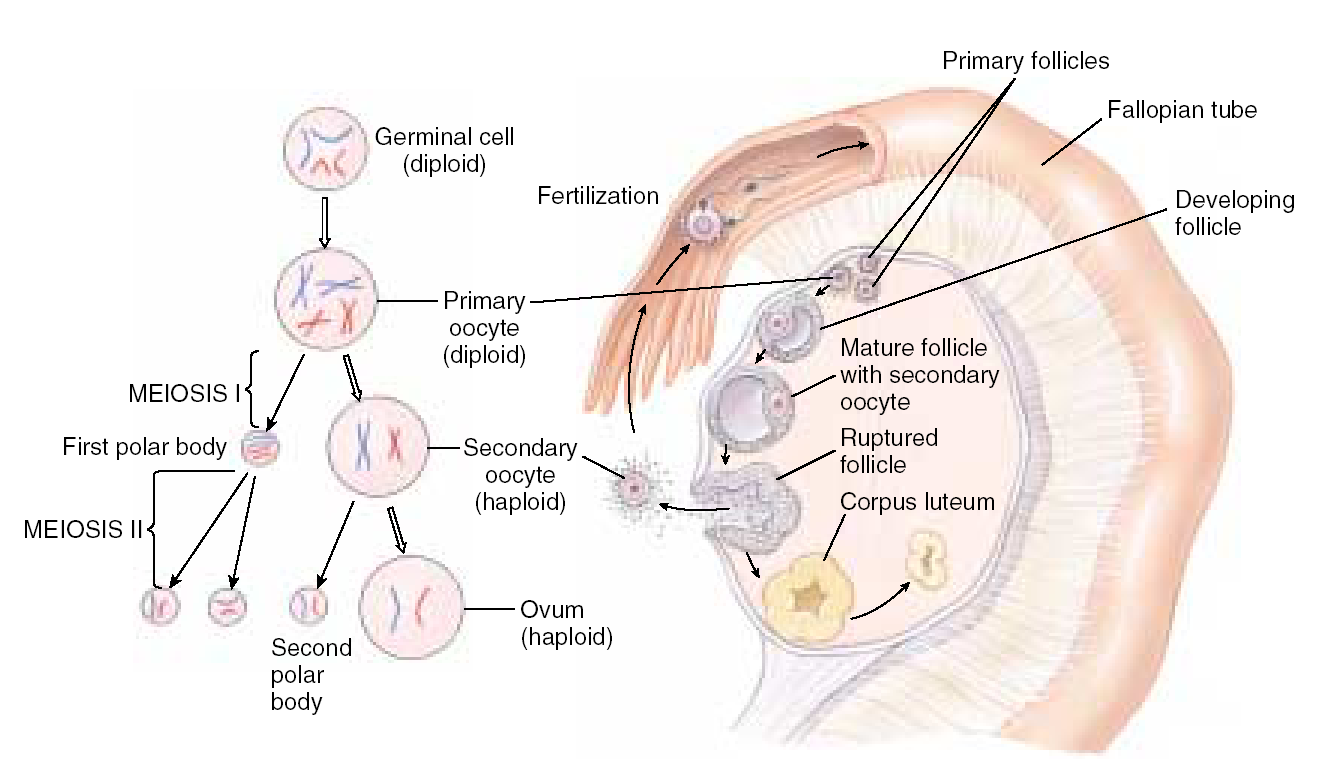 Рис. 6.4. Схема овогенез. млекопитающих и человека заканчивается еще до рождения. Сформировавшиеся к этому времени овоциты I порядка (2n4c) сохраняются в яичнике без изменения многие годы. С наступлением половой зрелости один раз в 28-32 дня один из овоцитов увеличивается в размерах (период роста (II), окружается фолликулярными клетками, обеспечивающими питание. Наступает период созревания (III). Под влиянием женских половых горомнов овоцит I порядка проходит редукционное деление мейоза. Образуется один овоцит II порядка и одно полярное тельце. Наступает второе мейотическое деление, которое в яичнике проходит до стадии метафазы. На этой стадии овоцит II порядка выходит из яичника в брюшную полость, а оттуда попадает в маточные трубы. Дальнейшее созревание не произойдет до тех пор, пока овоцит не объединится со сперматозоидом, который окажет стимулирующее воздействие. В маточных трубах овоцит II порядка уже после слияния со сперматозоидом завершает 2-е мейотическое деление и превращается в овотиду - зрелую яйцеклетку. Если оплодотворения не произойдет, овоцит II порядка погибает, слизистая матки отторгается и наступает менструация. Т.о. овогенез включает 3 периода: размножение, рост и созревание. Из одного овоцита I порядка образуется только 1 зрелая яйцеклетка и 3 полярных тельца. Морфология яйцеклетки.Яйцеклетка неподвижна, имеет шарообразную форму, содержит ядро, цитоплазму, все органоиды. В яйцеклетке содержатся питательные вещества, необходимые для зародыша (желток). По количеству желтка различают 3 типа яйцеклеток (рис. 6.5.): алецитальные (характерны для человека) и олиголецитальные (маложелтковые), мезолецитальные (среднежелтковые), полилецитальные (многожелтковые). По расположению желтка яйцеклетки бывают изолецитальными (желтка немного, он распределен 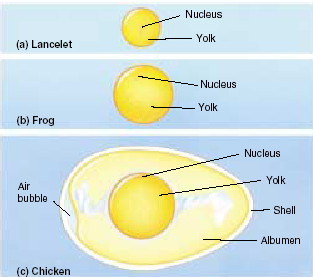 Рис. 6.5. Типы яйцеклеток. равномерно, такой тип характерен для низших хордовых, иглокожих, млекопитающих), телолецитальными (желтка много, он сосредоточен на одном из полюсов, характерен для птиц, земноводных, рептилий) и центролецитальными (желток находится в центре клетки вокруг ядра, встречается у насекомых). В овогенезе яйцеклетки приобретают полярность: формируются вегетативный и анимальный полюса, возникает различие состава участков цитоплазмы. Это явление называется овоплазматической сегрегацией. Основные виды размножения. Половое и бесполое размножение Размножение или репродукция – одно из основных свойств живого. Размножение подразумевает способность организмов производить себе подобных особей. В конечном итоге существование любого вида организмов поддерживается размножением. Известны две основные формы размножения – половое и бесполое. При бесполом размножении (рис. 6.7) организм возникает из соматических клеток и источником изменчивости могут быть случайные мутации. Бесполое размножение широко распространено среди растений, простейших и некоторых многоклеточных. Основные формы бесполого размножения – деление на два и множественное деление (шизогония). Деление на два сопровождается митозом, в результате образуются две идентичные особи. Такой тип деления распространен у простейших, встречается у амеб, эвглены зеленой. При множественном делении (шизогонии) ядро исходной клетки несколько раз делится митозом, и только потом происходит деление цитоплазмы. Шизогония встречается у малярийного плазмодия. При почковании новая особь образуется в виде выроста (почки) на родительской особи, а затем отделяется от нее, превращаясь в самостоятельный организм, идентичный родительскому. Почкование характерно для дрожжей, кишечнополостных. Фрагментация – это способ размножения фрагментами. Происходит при делении особи на 2 или больше частей, каждая из которых образует новую особь. С фрагментацией связана регенерация, т.е. способность восстанавливать целостный организм. Фрагментация встречается у плоских червей, морских звезд.  Рис. 6.7. Формы бесполого размножения Спорообразование. Спора –одна из стадий жизненного цикла, служащая для размножения. Спора состоит из клетки с ядром и цитоплазмой, покрытой оболочкой. Споры гаплоидны. Они способны разноситься на дальние расстояния, имеют приспособления для полета и для защиты от неблагоприятных условий. Спорообразование характерно для водорослей, грибов, папоротников, мхов. При вегетативном размножении от растения отделяется часть, способная развиваться в самостоятельное растение. Для вегетативного размножения могут служить специальные образования, имеющие запас питательных веществ: клубни (картофель), луковицы (лук, тюльпан), корни (ландыш, малина), усы (земляника). Для искусственного вегетативного размножения используют черенки (части стебля), отводки (ветки пригибают к земле и на них образуются корни – у малины, крыжовника), деление кустов (смородина, земляника), корневые отпрыски (малина), прививки. У организмов, размножающихся бесполым путем, новое поколение появляется из неспециализированных клеток тела. В основе их самовоспроизведения лежит митоз, обеспечивающий постоянство наследственного материала не только в ряду клеточных поколений, но и организмов. Примером бесполого размножения у многоклеточных является полиэмбриония (например, рождение однояйцевых близнецов у человека – в результате одного оплодотворения появляется 2 или более организмов). Половое размножение Хотя в процессе развития жизни бесполое размножение возникло первым, половое размножение существует на Земле более 3 млрд. лет. Половое размножение характеризуется наличием полового процесса, при котором происходит слияние гаплоидных (т.е. с одинарным набором хромосом) половых клеток (гамет). Гаметы образуются в результате особого вида деления клеток – мейоза. Половой процесс встречается у одноклеточных по типу конъюгации и копуляции. Конъюгация происходит у инфузорий при неблагоприятных условиях (рис. 6.8). Инфузории имеют 2 ядра: макронуклеус и микронуклеус. При конъюгации инфузории сближаются, между ними образуется цитоплазматический мостик. Макронуклеус, обеспечивающий обменные 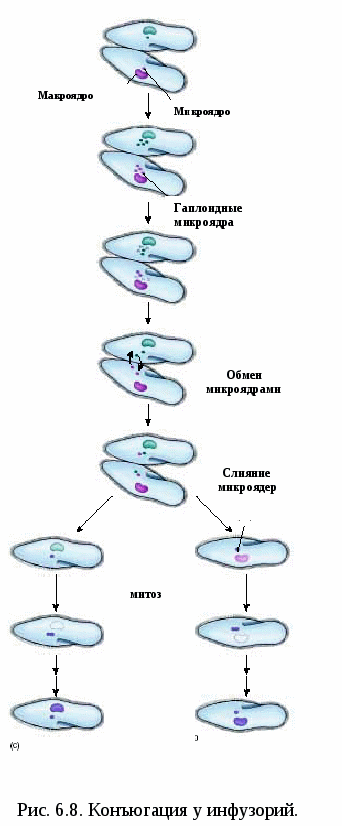 процессы, растворяется. Микронуклеус делится мейозом, в результате чего образуется 2 ядра с гаплоидными наборами хромосом (стационарное и мигрирующее). Мигрирующие ядра переходят через цитоплазматический мостик в цитоплазму партнера. Затем стационарное ядро 1 сливается с мигрирующим ядром 2, образуя, так называемый, синкарион. В синкарионе содержится диплоидный набор хромосом. После ряда сложных перестроек из синкариона формируются обычные микронуклеус и макронуклеус. После конъюгации инфузории расходятся, но, благодаря обмену ядрами, наследственная информация каждой особи изменяется, что приводит к появлению новых комбинаций и свойств. Копуляция – это половой процесс у одноклеточных организмов, при котором две особи приобретают половые различия, т.е. превращаются в гаплоидные гаметы, и полностью сливаются, образуя диплоидную зиготу. В процессе эволюции степень различия гамет нарастает. На I этапе различия между клетками отсутствуют (изогамия). Так, например, размножается жгутиконосец политома. Дальнейшее усложнение связано с дифференцировкой гамет на крупные и мелкие, т.е. появляется анизогамия (гетерогамия), т.е. неравный брак (например, Вольвокс). Такая форма анизогамии, когда гаметы резко различаются, называется оогамией. У многоклеточных при половом размножении имеет место лишь оогамия. Половое размножение многоклеточных Примером полового размножения многоклеточных является партеногенез – развитие из гамет без оплодотворения. Партеногенез встречается у дафний, а в неблагоприятных условиях - у пчел. В классическом варианте половой процесс включает период формирования половых клеток и оплодотворение (см. часть II). |